 2015-06-10
2015-06-10 2407
2407Соссюр, работая с зелеными растениями в темноте, обнаружил, что они выделяют СО2 даже в бескислородной среде. Л. Пастер нашел, что в темноте в отсутствие кислорода в растительных тканях наряду с выделением СО2 образуется спирт, т. е. идет спиртовое брожение. Он пришел к выводу, что в растительных тканях, так же как и у бактерий, возможно спиртовое брожение.
Немецкий физиолог Э. Ф. Пфлюгер (1875) показал, что лягушки в среде без кислорода некоторое время остаются живыми и при этом выделяют СО2. Пфлюгер назвал это дыхание интрамолекулярным, т. е. дыханием за счет внутримолекулярного окисления субстрата, и оно является начальным этапом нормального аэробного дыхания. Немецкий физиолог растений Б. Пфеффер распространил эту точку зрения на растительные организмы. Пфеффером и Пфлюгером были предложены два уравнения, описывающие механизм дыхания:
1) С6Н12О6 → 2С2Н5ОН + 2СО2
2) 2С2Н5ОН + 6О2 →4СО2 + 6Н2О
С6Н12О6 + 6О2 → 6СО2 + 6Н2О
На первом, анаэробном, этапе происходит спиртовое брожение, образуются две молекулы этанола и две молекулы СО2. Затем в присутствии кислорода спирт, взаимодействуя с ним, окисляется до СО2 и Н2О.
|
| |||
 | |||
| |||
В опытах Костычева и его сотрудников (1912 — 1928) было показано, что если растительные ткани кратковременно выдержать в бескислородной среде, а затем дать кислород, то наблюдается резкое усиление дыхания, т. е. в ходе анаэробной фазы накапливаются промежуточные продукты, которые в присутствии кислорода быстро используются. Ингибиторы, блокирующие брожение, например NaF, блокируют и аэробное дыхание. Костычев пришел к выводу о том, что промежуточным продуктом может быть уксусный альдегид. Благодаря работам немецкого биохимика К. Нейберга, Костычева и других стало очевидным, что дыхание и все виды брожения связаны между собой через пировиноградную кислоту (ПВК):
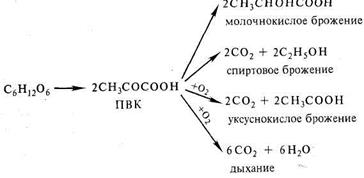
Глюкоза является стабильным соединением. Для того чтобы подвергнуться дыхательному распаду, она должна быть активирована. Значение анаэробного этапа дыхания и брожения состоит в преодолении химической инертности молекулы гексозы, т.е. в ее лабилизации и активации. Активация глюкозы происходит на первом, подготовительном, этапе гликолиза (см.гликолиз 4.1.2).
4. Основные пути диссимиляции углеводов.
Основными путями диссимиляции углеводов являются 1) гликолитический путь, 2) пентозофосфатный путь; 3) циклы ди- и трикарбоновых кислот.
Гликолитический путь, в основе которого лежит двукратное фосфорилирование гексозы, и ПФП с одним фосфорилированием глюкозы — не единственные пути окисления молекулы сахара. Некоторые организмы способны окислять и нефосфо-рилированную глюкозу. Этот путь прямого окисления сахаров обнаружен у некоторых бактерий, грибов и животных, а также у фотосинтезирующих морских водорослей. Ферментативное окисление глюкозы в глюконовую кислоту сопровождается выделением пероксида водорода, который затем разлагается каталазой или пероксидазой. Образовавшаяся глюконовая кислота может вовлекаться в дальнейший метаболизм после ее фосфорилирования через образование двух триоз − пировиноградной кислоты и 3-фосфоглицеринового альдегида, которые через ПВК могут окисляться в цикле Кребса.
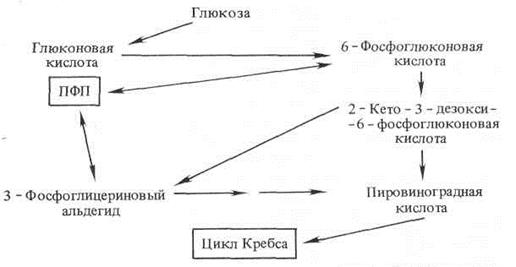 |
Если в процессе дыхания прямому окислению подвергаются и другие сахара, кроме глюкозы, то образуется целое семейство кислот, названных кислотами прямого (первичного) окисления сахаров.
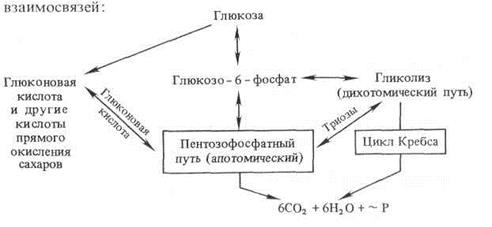 |
Дыхательные циклы − гликолиз и цикл ди- и трикарбоновых кислот, ПФП и прямое окисление сахаров − система взаимосвязанных процессов. Ниже представлена схема этих взаимосвязей:
Связь между гликолизом и ПФП осуществляется через глюконовую кислоту и фосфотриозы. В клетке гликолиз и ПФП пространственно не отделены друг от друга. Эти процессы протекают в растворимой в растворимой части цитоплазмы, в пропластидах и хлоропластах. Они имеют общие субстраты — глюкозо-6-фосфат, фруктозо-6-фосфат и 3-фосфоглицериновый альдегид. В норме доля пентозофосфатного цикла в общем дыхательном обмене составляет 10−40% и варьирует в зависимости от типа ткани и ее функционального состояния. В анаэробных условиях гликолиз доминирует над ПФП. Однако в хлоропластах активность окислительного апотомического пути намного выше по сравнению с гликолизом. В цитоплазме большая часть продуктов ПФП метаболизируется через гликолиз.
Активность ПФП увеличивается при неблагоприятных условиях: засухе, калийном голодании, инфекции, затенении, засолении, при старении.

