 2015-06-16
2015-06-16 4175
4175Регулирование конечным продуктом активности аллостерического фермента определенного биосинтетического пути обеспечивает мгновенную реакцию, приводящую к изменению выхода этого продукта. Если последний оказывается ненужным, отпадает надобность и в ферментах, участвующих в его синтезе. Проявлением максимальной экономичности клеточного метаболизма служат выработанные клеткой механизмы, регулирующие ее ферментный состав. Очевидна целесообразность синтеза только тех ферментов, которые необходимы в конкретных условиях. Показано, что у прокариот в одних условиях фермент может содержаться в количестве не более 1 — 2 молекул, в других — составлять несколько процентов от клеточной массы.
Количество определенного фермента в клетке может регулироваться на нескольких уровнях: на этапе транскрипции, трансляции, а также в процессе сборки и разрушения ферментного белка (см. рис. 28). В иерархии регуляторных воздействий наиболее сложный механизм, контролирующий количество ферментов в клетке, связан с процессом транскрипции. Специфические химические сигналы могут инициировать или блокировать транскрипцию определенного участка ДНК в иРНК. В случае индукции образованная иРНК участвует в определенной последовательности реакций, называемой трансляцией и заканчивающейся синтезом полипептидных цепей. Регуляция белкового синтеза на уровне трансляции может осуществляться на любом из ее этапов, например на этапе инициации, элонгации и др. Не исключена также возможность изменения времени жизни иРНК. под воздействием разных эффекторов, в том числе конечных продуктов метаболических путей. Хотя механизмы регуляции синтеза белка на уровне трансляции еще точно не установлены, ясно, что на этом этапе имеются широкие возможности для регуляции скорости синтеза различных белков.
Известно, что фермент может выполнять метаболическую функцию после приобретения соответствующей структуры. Скорость образования структур высшего порядка также находится под контролем определенных молекул. Таким образом, контроль на уровне сборки функционально активного фермента может играть существенную роль в метаболической регуляции. Наконец, скорость разрушения фермента под воздействием специфических метаболических сигналов будет также определять его концентрацию в клетке.
Регуляция синтеза ферментов на этапе транскрипции основана на том, что "считывание" бактериальных генов происходит избирательно и скорость образования копий соответствующих иРНК (а отсюда и дальнейшая их трансляция в белки) находится под сложным контрольным механизмом. Скорость синтеза ферментов, определяемая этой стадией, может меняться в разной степени. По данному признаку все ферменты делятся на два класса. Ферменты, синтез которых в растущей клетке происходит с постоянной скоростью в результате постоянного транскрибирования соответствующих генов и, следовательно, они присутствуют в клетке в более или менее постоянной концентрации, называются конститутивными. К ним относятся, например, гликолитические ферменты. Метаболические пути, функционирующие с участием конститутивных ферментов, контролируются посредством других регуляторных воздействий, например аллостерического ингибирования.
Кроме этого в бактериальных клетках имеются ферменты, количества которых могут резко меняться в зависимости от состава питательных веществ среды. Это происходит в результате того, что гены, детерминирующие эти ферменты, включаются или выключаются по мере надобности. Их называют индуцибельными. При отсутствии в среде субстратов этих ферментов последние содержатся в клетке в следовых количествах. Если в среду добавить вещество, служащее субстратом определенного фермента, происходит быстрый синтез этого фермента в клетке, т. е. имеет место индукция синтеза фермента. Если же в питательной среде в готовом виде содержится вещество, являющееся конечным продуктом какого-либо биосинтетического пути, происходит быстрое прекращение синтеза ферментов этого пути. Это явление получило название репрессии конечным продуктом. Ферменты, синтез которых подавляется конечным продуктом, могут быть дерепрессированы, т. е. скорость их синтеза превысит обычную, если концентрация конечного продукта упадет до очень низкого уровня. Дерепрессия этих ферментов аналогична явлению индукции.
Репрессия конечным продуктом. Все биосинтетические пути находятся под контролем механизма репрессии конечным продуктом. Точно так же образование большинства анаболических ферментов регулируется путем репрессии их синтеза. Репрессия осуществляется особыми присутствующими в клетке веществами — репрессорами. Факторами, модифицирующими активность репрессоров, могут быть конечные продукты биосинтетических путей, а также промежуточные продукты некоторых катаболических или амфиболических путей.
Репрессия может быть координированной, т. е. синтез каждого фермента данного пути в одинаковой степени подавляется конечным продуктом. Часто синтез ферментов одного пути репрессируется в разной степени. В разветвленных биосинтетических путях механизмы репрессии могут быть модифицированы (как и механизмы ингибирования), чтобы лучше обеспечить регуляцию нескольких конечных продуктов из общего исходного субстрата. Синтез многих ферментов в таких путях репрессируется только при совместном действии всех конечных продуктов. Если реакция на общем участке разветвленного пути катализируется изоферментами, синтез каждого из них находится под контролем "своего" конечного продукта (см. рис. 31).
Механизм репрессии конечным продуктом на уровне транскрипции стал проясняться с 50-х гг. Большой вклад в это внесли работы Ф. Жакоба и Ж. Моно. Было показано, что наряду со структурными генами, кодирующими синтез ферментов, в бактериальном геноме существуют специальные регуляторные гены. Один из них — ген-регулятор (ген R), функция которого заключается в регуляции процесса транскрипции структурного гена (или генов). Ген-регулятор кодирует синтез специфического аллостерического белка-репрессора, имеющего два центра связывания: один узнает определенную последовательность нуклеотидов на участке ДНК, называемом оператором (ген О), другой — взаимодействует с эффектором. Ген-оператор расположен рядом со структурным геном (генами) и служит местом связывания репрессора. В отличие от операторных генов гены-регуляторы расположены на некотором расстоянии от структурных генов (продукты регулярных генов — репрессоры являются свободно диффундирующими белковыми молекулами).
Часто структурные гены, относящиеся к одному биохимическому пути, объединены в группу, составляющую вместе с оператором единицу транскрипции и регуляции — оперон. Все структурные гены, объединенные в оперон, имеют один операторный участок, локализованной на краю оперона, и координирование регулируются одним репрессором. Оперон представляет собой весьма рациональную и эффективную систему регуляции метаболического пути.
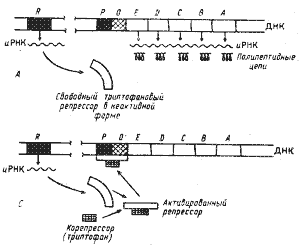 |
| Рис. 32. Триптофановый оперон Е. coli и механизм репрессии конечным продуктом. А — продукт гена-регулятора (R) — неактивная форма репрессора, неспособная связываться с оператором (О); промоторный участок (Р) открыт: происходит транскрипция структурных генов (Е, D, С, В, А). Б — в присутствии корепрессора образуется активный комплекс корепрессор — репрессор, связывающийся с оператором и закрывающий промотор; транскрипции не происходит (по Lehninger, 1974) |
Процесс транскрипции начинается с прикрепления РНК-полимеразы, катализирующей синтез иРНК, к определенному участку ДНК, называемому промотором (Р). Когда молекула репрессора "садится" на операторный участок, она "закрывает" промотор, тем самым препятствуя связыванию с ним РНК-полимеразы и началу транскрипции. У прокариот пять генов, кодирующих синтез ферментов триптофанового пути, образуют оперон (рис. 32). Ген-регулятор обеспечивает синтез аллостерического белка — триптофанового репрессора, не активного в свободном состоянии. Последний в таком виде не связывается с операторным участком и, следовательно, не может препятствовать началу транскрипции. Когда конечный продукт метаболического пути (триптофан) накапливается выше определенного уровня, он взаимодействует с репрессором и активирует его.
Активированный репрессор присоединяется к операторному участку и подавляет транскрипцию триптофанового оперона. Таким образом, триптофан является корепрессором.
Индукция синтеза ферментов. В большинстве случаев регуляция путем индукции характерна для катаболических путей, где в качестве индукторов выступают обычно субстраты этих путей. Классический пример индуцибельного фермента — (3-галактозидаза Е. coli. Оказалось, что если клетки Е. coli выращивать в среде, содержащей глюкозу, то они не могут использовать лактозу. Если такие клетки поместить в среду, где лактоза — единственный источник углерода, после некоторого периода в них происходит интенсивный синтез фермента (3-галактозидазы, катализирующего гидролиз лактозы на D-глюкозу и D-галактозу. С помощью этого фермента Е. coli может теперь использовать лактозу в качестве единственного источника углерода. Если затем клетки, растущие на среде с лактозой, перенести на среду с глюкозой, синтез (3-галактозидазы прекращается.
Изучение индукции (3-галактозидазы у Е. coli позволило установить, что рост клеток на среде с лактозой происходит не в результате отбора мутантов, у которых способность использовать лактозу есть следствие мутации. Способностью синтезировать этот фермент обладают все клетки. Было также показано, что в процессе индукции происходит не активирование уже имеющегося в клетках фермента (З-галактозидазы, а его синтез de novo из аминокислот.
Индуцированный синтез ферментов у микроорганизмов был описан в 30-х гг., но механизм этого процесса долгое время оставался непонятен. Индуцированный синтез ферментов лежит в основе широко известного явления адаптации организмов к различным условиям. Успехи, достигнутые в расшифровке механизмов регуляции клеточного метаболизма, позволили объяснить природу этого явления, его механизм и роль в клетке.
Лактозный оперон Е. coli, состоящий из трех структурных генов, промотора и оператора, был первой ферментной системой, на которой Ж. Моно и Ф. Жакоб изучали механизм индукции синтеза ферментов (рис. 33). В отсутствие лактозы молекула репрессора, активная в свободном состоянии, связывается с оператором и подавляет транскрипцию структурных генов. Когда в клетку попадает лактоза, она связывается с репрессором, в результате образуется неактивный комплекс репрессора с индуктором, который не может взаимодействовать с оператором и, следовательно, препятствовать транскрипции структурных генов. В результате индуцируется синтез ферментов катаболизма лактозы. При удалении из клетки индуктора репрессор снова переходит в активное свободное состояние, связывается с оператором, что приводит к прекращению синтеза соответствующих ферментов.
Катаболитная репрессия. Кроме репрессии конечным продуктом, характерной для анаболических путей, описан тип репрессии, называемой катаболитной и заключающейся в том, что быстро используемые клеткой источники энергии способны подавлять синтез ферментов других путей катаболизма, участвующих в метаболизировании сравнительно медленно используемых источников энергии. Катаболитную репрессию можно рассматривать как приспособление клетки к использованию в первую очередь наиболее легко доступных источников энергии. В присутствии такого источника энергии потребление других субстратов, менее "удобных" для клетки, временно приостанавливается, и пути катаболизирования этих субстратов временно выключаются.
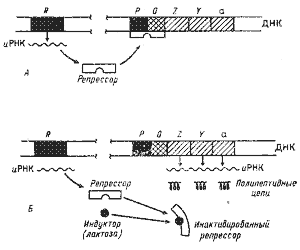 |
| Рис. 33. Индукция синтеза ферментов катаболизма лактозы у Е. coli. А — ген-регулятор (R) образует репрессорный белок, связывающийся с оператором (О) и закрывающий промотор (Р); транскрипции структурных генов (z, у, а) не происходит; Б — в присутствии индуктора образуется неактивный репрессор, теряющий способность связываться с оператором; промотор открыт, происходит транскрипция |
Выше уже отмечалось, что если в среде для выращивания Е. coli одновременно содержатся глюкоза и лактоза, сначала используется глюкоза. Несмотря на присутствие индуктора лактозного оперона, ферменты, участвующие в катаболизме лактозы, не синтезируются. Транскрипция генов лактозного оперона начинается, когда концентрация глюкозы в среде становится низкой. Таким образом, глюкоза препятствует синтезу ферментов лактозного оперона.
Как это осуществляется? Изучение механизма катаболитной репрессии обнаружило, что этот тип регуляции тесно связан с внутриклеточным уровнем циклического АМФ (цАМФ), который в этом процессе функционирует в качестве эффектора. Он образует комплекс с аллостерическим белком — катаболитным активатором, не активным в свободном состоянии. Этот комплекс, присоединившись к определенному участку на промоторе, обеспечивает возможность связывания РНК-полимеразы с промотором и инициацию транскрипции. Количество образующегося комплекса определяется концентрацией цАМФ, которая уменьшается при увеличении содержания глюкозы в среде. Таким образом, глюкоза вызывает изменение внутриклеточной концентрации цАМФ. Это соединение обнаружено в клетках всех прокариот. Его единственная функция — регуляторная. Циклический АМФ образуется из АТФ в реакции, катализируемой аденилатциклазой, связанной с ЦПМ:
АТФ цАМФ + пирофосфат.
Аденилатциклаза обладает высокой активностью, если компоненты системы транспорта глюкозы в клетку фосфорилированы. Это происходит в отсутствие глюкозы, которую необходимо транспортировать. Таким образом, активность аденилатциклазы возрастает при уменьшении концентрации глюкозы в среде. Последнее приводит к повышению образования цАМФ и в конечном итоге к индукции синтеза ферментов катаболизма лактозы. Наоборот, при высокой концентрации глюкозы в среде система ее транспорта находится в дефосфорилированном состоянии, следствием чего является уменьшение активности аденилатциклазы и соответственно количества цАМФ. Таким способом глюкоза через систему своего транспорта регулирует концентрацию цАМФ в клетке. Поскольку катаболизм глюкозы связан с образованием метаболической энергии и запасанием ее в молекулах АТФ, через глюкозу в клетке связаны пулы АТФ и цАМФ: при увеличении количества АТФ уменьшается количество цАМФ и наоборот.
Особенностью всех ферментных систем, находящихся под контролем катаболитной репрессии, является участие в их индукции универсального комплекса, состоящего из белкового катаболитного активатора и цАМФ.

