 2015-06-28
2015-06-28 3215
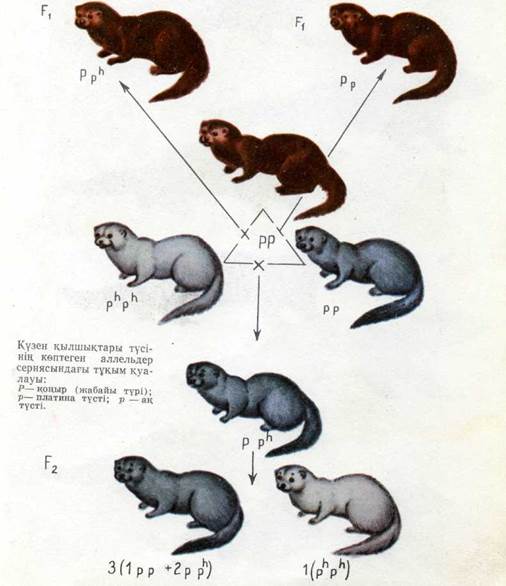
3215Гетерозис туралы ұғым. Жануарлар мен өсімдіктер селекциясында гибридтік буданды, ңуаттылық қүбылысы немесе гетерозис айрықша орын алады, ол мынадайлардан тұрады. Жануарлар түқымдары мен өсімдіктер сорттарының алуан түрлерін, нәсілдерін, сондай-ақ инбредті линияларды шағылыстыру кезін-де, Ғ\ будандары бастапқы ата-ана организмдерден өсу энер-гиясы, қоректік заттарды жақсы пайдалануы, әр түрлі ауруларға төзімділігі және т. б. женінен көбіне асып түседі. Ғ\ будандарын езара шағылыстыру бұл нәтиженің келесі үрпақтарда тоқталуына әкеледі.
Дарвиннің пікірі бойынша гетерозис — түрлер эволюция-сында шағылысудың биологиялық пайдалы себептердің бірі болып табылады.- Айқас үрықтану табиғи сүрыптаумен сақта-лып отырады, өйткені айқас үрықтану гибридтік қуаттылықты қолдаудың механизмі болып табылады. Жүгерінің линия аралық будандары. Осы ғасырымыздың басында жүгеріде инбредті линиялардың арасындағы шағылысуды зерттеу жүйелі түрде жүргізіле бастады. Бүл жағдайда кейбір линияларды шағылыстырудан бастапқы линиялар мен сорттарға қарағанда өте өнімді будан өсімдіктер алынатынын Г. Шелл дәлелдеді. 11-таблицада өсімдіктер өздігінен тозаңдан-ғанда инбредті линиялардың төмен өнім беретіндігін, Ғі-де өнім-нің недәуір артатындығын және оның Ғ2-ле төмендеуін көрсете-тін деректер келтірілген.
Қазір буданды тұқым себу жүгері өндірудегі негізгі тәсіл-дердін, біріне айналды. Буданды тұқым алу үшін әуелі 5—7 жыл бойына сол ауданның климатының талаптарына сай келетін таңдаулы сорттардың көптеген инбредті линиялары жасалады. Линиялардың көпшілік бөлігі (99°/о-ке жуығы) әр түрлі жа-ғымсыз қасиеттеріне қарай бракқа шығарылады.
11 -таблица
Жүгеріні шағылыстырудың эр түрлі типтеріндегі өсімдіктердің қуаттылық көрсеткіштері

Инбредті линияларды жасау — гетерозисті формалар алу үшін жұмыстың маңызды кезеңі. Линиялар ішіидегі особьтар-да ұқсас генотиптер болады және іс жүзінде гомозиготалы бо-лып табылады. Сондықтан мұндай линиялардың шағылысуы генотиптері бойынша бірдей гетерозиготалы будандар береді.
Көп мөлшерде инбредті линияларды жасаған соң, олардың арасында шағылыстыру жүргізуге кіріседі. Бірінші үрпақтың линиялар аралыщ будандарын гетерозистің эффектісі бойынша бағалайды, жақсы комбинациялар беретін линияларды сүрып-тайды, одан сон, олардан буданды тұқымдар өндіру үшін үлкен көлемде көбейтеді. Шағылысу кезінде жоғары тиімділікпен гетерсззис беретін екі линияны табу үшіи, бірнеше мыңдаған будандар комбинацияларын тексеру қажет.
Шағылысу кезінде ең үлкен тиімділікпен гетерозис беретін бастапқы линиялардан өндіріс мақсаттары үшін буданды тү-қымдарды алу үшін аналық және аталық формаларды қатар-ластыра себеді. Олардың арасындағы тозаңдануды қамтама-сыз ету үшін цитоплазмалық аталық стерильділікті пайдалңна-тын буданды түқымдарды өндіру схемасы жасалған (10-тарауды қараңыз), бұл аналық линиялар өсімдіктерінен шашақ бас гүл-дерін жұлу қажетті болмағандықтан, еңбек шығынын недәуір қысқартуға мүмкіндік берді. Жүгерінің қарапайым аралық бу-дандарын осылай а лады. Бүл әдіс айқас тозаңданатын әр түрлі өсімдіктер будандарының тұқым шаруашылығы үшін ортақ болады.
. Қазіргі уақытта қосарланған линия аралық будандар кеңінен пайдаланылады. Оларды гетерозис көрсететін екі қарапайым буданды шағылыстыру жолымен алады (104-сурет). Неғұрлым өнімді қосарланған будандарды алу үшін карапайым будандарды сүрыптау селекцияның маңызды кезеңі болып табылады. Әр түрлі сорттардан шығатын линияларды шағылыстыру жақсы
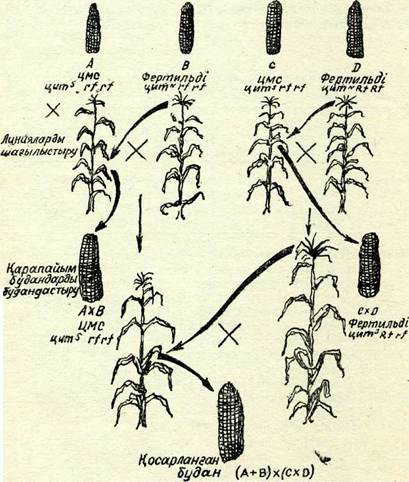
104-сурет. Цитоплазмалық аталық стерильділігін пайдалану арқылы жүгері-нің қосарланған буданын алу схемасы. Белгілерін 57-суреттен қараңыз.
нәтижелер береді. Мәселен, егер бір қарапайым будан АУ(В екі сорттарының инбредті линияларын шағылыстырудан, ал басқа-сы С~ХД сорттарының линияларын шағылыстырудан алынса, досарланған будан (АхВ)Х(СхД) бір сорт линияларынан шы-ғарылған қарапайым будандарды шағылыстырудан алынған қосарланған гибридтен гөрі гетерозисті жиі береді:
(ЛХ^і) X (А2ХАъ) немесе (ВХ^і) X (ВяХЗД.
Жүгеріден будандарды алу туралы айтылғанның бәрі ай-қас тозаңданушы басқа өсімдіктерге де (пияз, қызылша және т. б.) қатысты болады. Әр түрлі өсімдіктердің линия аралық гибридтері бастапқы сорттардан өнімділігі бойынша 40—8О°/о-ке артық өнім береді.
Жануарлардан да гибридтерді осыған ұқсас жолмен алады. Казіргі уақытта құс шаруашылығы мен шошқа шаруашылы-ғында бір немесе әр түрлі тұқымдардан шығатын инбредті линияларды шағылыстыру кеңінен пайдаланылуда.
Инбредті линиялар әр уақытта сортпен салыстырғанда темен көрсеткішке ие болатындықтан, линия аралық будан бастапқы линиялардан ғана емес, сондай-ақ сол линиялар шыққан сорттар мен тұқымдардан асып түскен жағдайда ғана гетерозис болуы туралы айтуға болады.
Гетерозистің көрінуі. Буданда гетерозистіц көрінуі, сондай-ақ цитоплазманың қасиеттеріне байланысты, мүның нәтижесін-де реципрокты шағылыстыру әр түрлі эффект береді. Мәселен, $ жылқы X & есекті шағылыстырудан гетерозисі жоғары үзақ жасайтын, төзІмді және күшті гибрид —■ қашыр алынады. Реципрокты комбинация гетерозисі мүлде жоқ қащыр (лошак) береді.
Буданды организм дамитын орта факторлары гетерозистің көрінуіне күшті эсер етеді. Гибридтердегі гетерозис бейімдел-гіш реакциялардың барынша үлкен нормасымен қамтамасыз етіледі деген болжамды тіпті Ч. Дарвин айтқан болатын. Соңғы жылдардағы зерттеулер Ғ\ гибридтерінде инбредті линиялар-мен салыстырғанда әр түрлі метаболиттер, есу заттары, энзим-дер және т. б. саны көп болатындығын көрсетті,
Гетерозистің мүмкін механизмдері. Қазір көптеген тендер бойынша гетерозиготалық күйде болудан, қолайлы доминант-ты геидердің өзара әрекеттесуінен, ңосарланған доминанттылықтан тыс болудан деген сияқты гетерозистің пайда болуын түсін-діруге тырысатын бірнеше гипотезалар бар.
Бұрын айтылғандай, гомозиготалы инбредті линияларды ша-ғылыстырғанда бірінші ұрпақтың будандары көптеген тендер бойынша гетерозиготалы болады. Бұл жағдайда зиянды рецессивті мутантты аллельдердің әрекеті екі ата-ананың доминантты аллельдерімен басып тасталынады. Схемамен алғанда мүны былай түсінуге болады: бір инбредті линия гомозиготалы күйде бір геннің (ааВВ) рецессивті аллеліне, ал екіншісі басқа геняің (ААЬЬ) аллеліне ие болады. Бүл рецессивті аллельдердің әр-қайсысы гомозиготалы күйде инбредті линияныц тіршілік қабілетін төмендететінбелгілі бір кемшілікті айқындайды, ааВВ X X A A bb линияларын ціағылыстырғакдд була«.^а.р^г. %«і тг\ш\я (АаВЬ) доминантты аллельдері бірігеді. Ғ\ будандары бұл жағ-дайда көрсетілген тендер бойынша тек гетерозисті ғана емес, сондай-ақ біркелкілікті де көрсетеді. Ғ2-де гетерозиготалы куй-де екі доминантты гендері бар особьтар (АаВЬ) саны Уі6 ғана болады, сондықтан особьтардың бәрі гетерозисті болмайды. Келесі ұрпақтарда гетерозиготалар саны кемиді, ал гомозигота-лар саны артады. Осы себептердің нәтижесінде гетерозис келе-сі ұрпақтарда тоқталады. Алайда барлық линия аралық гетерозиготалы будандар гетерозисті керсете бермейді, яғни гете-розиготалылық әр уақытта гетерозиспен байланысты болмайды..
Екінші гипотеза мына фактіден шығады, доминантты аллель-дер (жабайы типтің) рецессивті аллельдерге қарағанда көбінесе қолайлы эсер көрсетеді. Сондықтан будаи комбинациясын-да доминантты аллельдерді біріктіру гетерозисті тезірек қамтамзсыз етеді. Басқаша айтқанда, бұл гипотеза доминантты аллельдердің комплементті әрекеттесу тиімділігінін қарапайым қосындысы туралы түсініктен шығады. Бұл гипотеза да біраз фактілерге қайшы келеді. Бұл гипотезаға сәйкес, инбридинг нәтижесінде доминантты гендердің гомозиготалы күйдегі жиынтығы бар гетерозисті формалардың пайда болуын күту қажет, алайда ол байқалмайды.
Үшінші гипотеза аллельдердің гетерозиготалы күйі гомози-готалардан (АА<_Аа^>аа) артық болады дегеннен шығадьь Мұнда гетерозиготада аллельдердің жабайы және мутантты типінің үйлесуі доминанты гендердің әрекетін күшейтеді және осыған байланысты синтезі осы геидермен бақыланатын ерекше заттардың барынша жиналуына жағдай жасайды деп жора-малдауға болады. Гетерозисті осылайша түсіндіруді доминант-тықтан тыс гипотеза деп атайды. Бұл аллель аралық комплементациямен байланысты болуы мүмкін (17-тарауды қараңыз).
Жуырда В. А. Струнников гипотеза ұсынды, бұл гипотезаға сәйкес линияда инбридинг пен гомозиготизация нәтижесінде гомозиготалы мутантты аллельдердің зиянды әрекетіне кедергі жасайтын гендердің компенсаторлы комплекс! жиналады. Әр түрлі инбредті линияларды шағылыстырғанда олардың әрқай-сысы буданға осы комплексті енгізеді, ал зиянды мутациялар гетерозиготалы күйге ауысады, мүның нәтижесінде гетерозис байқалады.
Бүл гипотезалардың біреуін де бірдеи-бір дұрыс гипотеза деп есептеуге болмайды. Шамасы, бүл гипотезалар қарастырған механизмдердің әрқайсысы будандық қуаттылықты анықтау-да белгілі бір роль атқарады.
Гетерозисті бекіту жолдары. Селекцияда гетерозисті пай-даланудың негізгі міндеттерінің бірі оны бекіту, яғни гибридті өндіру процесінде гетерозистік эффектіні сақтау болып табыла-ды. Бүл міндеттерді шешу бірнеше аспектілерде ойластырылады. Гетерозисті бекітуді гибридтік организмді қалыпты жыныстық көбеюден апомиксистік көбеюге ауыстыру жолымен жүзеге асыруға болады (3-тарауды қараңыз), бұл кейбір өсімдіктер үшіи мүмкін. Диплоидты апомиксисті пайдалану кез келген ең күрделі деген гендердің гетерозиготалы жүйесін сақтау үшІн перспектива ашады. Бүл жағдайда аса үздік қасиеттері бар бір өсімдік сортқа бастама бере алады.
Вегетативті көбейетін өсімдіктерде, жыныстық жолмен алын-ған бағалы гибридтік комбинацияларды, вегетативті кебеюдің езімен — қалемше, ұластыру, түйнек және т. б. (5-тарауды қара) сақтайды. Қез келген мәдени өсімдіктер ушін жақсы буданды бір өсімдік вегетативті көбеюге ауысқанда, көптеген жеміс тү-қымдастар сияқты, өзінің бағалы қасиеттерін шексіз ұрпақтар бойына сақтайтын, санын кез келген мөлшерге кебейте алатын сортқа бастама бере алады.
Гетерозис! байқалатын диплоидты буданды полиплоидтық күйге ауыстыру жолымен де гетерозисті бекітуге мүмкін болады. Бүл жағдайда гендердің гетерозиготалы комбинацияларын бірқатар үрпақтар бойына сақтау мүмкіндігі недәуір жоғары (13-тарауды қараңыз).
Жануарларда гетерозисті бекітудің мүндай барлық жолда-ры қолданылмайды, сондықтан алмастырып шағылыстыру, яғ-ни будандарды кезекпен бастапқы форманың бірде бірімен екіншіде басқасымен шағылыстыру қолданылады. * -к А Комбинативтік өзгергіштік пен популяциялар генетикасы-ның заңдылықтарын білу — селекцияда шағылыстырудың әр түрлі әдістерін жаңа тұқымдар мен сорттар шығару үшін де, сондай-ақ тіршілік қабілеттері жоғары және енімді гетерозистік будандарды алу үшін де тиімді пайдалануға мүмкіндік береді.
33-тарау. ТҰҚЫМ ҚУАЛАУ
Сандық белгілер езгергіштігінің келемі өзара эсер ететін гендердің санына байланысты болады (7-тарауды қараңыз). Полигенді белгілердің қисық түрде өзгеруі 34-суретТе кескін-делген.
Модификациялық өзгергіштікті баяндау кезінде варианттар-дың қалыпты қисық таралуы келтірілген (73-сурет). Бұл қисық таралу толық ұқсас келеді.
Ал енді тұқым немесе сорттағы қандай да болмасын бір ша-руашылық пайдалы белгінің езгергіштігін кәз алдарыңа келті-ріңдер. Әлгінде ғана келтірілгенге ұқсас алынған қисық таралу немен камтамасыз етіледі? Үқсас генотиптердің особьтарында модификациялық езгергіштіктің бар болуынан ба немесе осы особьтардың (егер белгі полигенді болса) генотіштерішн, сан алуандығынан ба? Генотиптік өзгергіштікті фенотиптік өзгер-гіштіктен айыруға болатын бірден-бір әдіс математикалық әдіс иолып таоылады, оның көмегімен белгілердің тұқым қуалауы зерттеледі.
Тұқым қуалау деп популяциядағы зерттелетін белгінің өзгер-гіштігінің туқым қуалай қамтамасыз етілуі, ал тұқым қуалау дәрежесі деп генотиптік гетерогенділікпен қамтамасыз етілетін белгінің фенотиптік алуан түрлілігінің үлесі түсініледі. Тұқым қуалау сөздің кең мағынасында — бұл генотиптік өзгергіштік-тің фенотиптік өзгергіштікке қатынасы.
1. ТҰҚЫМ ҚУАЛАУ КОЭФФИЦИЕНТІ
Бұл қатынасты арнаулы математикалық әдістермен анық-тауға, яғни процентпен (1-ден 100%-ке дейін) немесе бірліктің улесімен (0-ден 1,0-ге дейін) белгіленетін тұқым қуалау коэ-фициентімен (һ2) есептеуге болады.
/i2=100% бақыланатын барлық особьтардың сан алуандығы олардың генотиптерінің айырмашылығымен (генотиптік өзгер-гіштікпен) қамтамасыз етілгенін білдіреді. Һ2 = О°/О. Әр түрлі особьтар генотиптерінің толық ұқсастығы жағдайында фенотип-тік сан алуандылықтың болуын, яғни модификациялық өзгер-гіштіктің орын алатынын білдіреді. /г2-нің аралық мәні популя-цияға генотиптікте, сол сияқты фенотиптікте өзгергіштіктін, бар екендігін көрсетеді.
Төменде ауыл шаруашылық жануарларыньщ әр турлі белгілерінің тұқым қуалау коэффициентінің (һ2) мәні келтірілген.
Кейбір белгілер үшіи тұқым қуалау коэффициентінің өзгер-мелілігіиің үлкендігі ең алдымен бул белгілер бойынша популя-циялардың табиғи айырмашылықтарының болуымен түсіндірі-леді. Әр түрлі популяциялардың (тұқым, табын) тұқым қуалай-тын гетерогенділігі мен гетерозиготалылығы оларды өсірудің бұрынғы тарихының нәтижесі болып табылады: инбридингтІ қолдану дәрежесі мен сипаты, популяция құрылымының қалып-тасуы және линияларға дифференциациялануы, сұрыптау және басқа популяциялармен шағылыстыру сипаты.
Мүйізді ірі қара
Сүттің сауылуы— 0—67%
Сүттегі майдыд мөліиері0—78%
Өсімталдығы0—18%
Туған кездегі тірі массасы 26—72%
Қой
Жүн қырку30—60%
Туған кездегі тірі массасы30—40%
Өсімталдығы '10—20%
Тауық
Жұмыртқа салғыштығы11—35%
Жумыртқа массасы30—70%
Тірі массасы 30—52%
Морфологиялық белгілер биологиялық бейімделушІлікке: өсімталдылық және тіршілікке қабілеттілікке байл.аныстыгі5ел~-гілермен салыстырғанда өте жоғары тұқым қуалауға ие. Өсім-талдылық пеи тіршілікке қабілеттіліктің тұқым қуалау коэф-фициеңтінің төмен болуын, бұл белгілерді анықтайтын тендер бойынша популяцияның гетерогенділігінің аздығымен түсіндіру-ге болады, бұл, әрине, эволюциялық тиімді.
2. СЕЛЕКЦИЯ ҮШІН ТҰҚЫМ ҚУАЛАУ КОЭФФИЦИЕНТІНІҢ МАҢЫЗЫ
Популяцияның тұқым қуалайтын гетерогенділігі сұрыптау-ға тиімді басты алғы шарттардың бірі болуға жақын. Сондық-тан нақты табыидағы немесе популяциядағы белгілердің тұқым қуалауын білу — организмдердің өнім бергіштігі мен тұқымдық бағалылығын арттыру жолдарын іздеуші селекционер үшін мін-детті болып табылады. Егер популяция генотиптері ұқсас особь-тардан тұрса, онда мұндай популяциядағы сүрыптаудың перспективасыз болатындығы күмәнсыз және бақыланатын бар-лық фенотиптік өзгергіштік мүндай популяцияда ортаның эсер етуімен қамтамасыз етіледі.
Түқым қуалауды білу.селекцияның жоспарланған тиімділігін анықтау үшін өте манызды. Мәселен, еріксіздікте 60 жылға жуық өсірілген күміс түстес қара түлкілерді сүрыптау арқылы олардың көбею маусымын жылжытудағы барлық әрекет нәти-жесіз болды. Д. К. Беляевтің анықтағанындай, бүл белгінің тү-қым қуалауы өте темен (1—2%) екені айқындалды, соның сал-дарынан бүл бойынша сүрыптау нәтижесіз болып табылады. Баска мысал келтірейік. Қырқылған жүнінің тұқым қуалауы бойынша елеулі айырмашылықтары бар биязы жүнді қойлардың екі тобы алынды: түқым қуалау бірінші топта 15,4%-ке, ал екін-ші топта—1,2%-ке тең. Осы екі топтағы сүрыптаудың тиімділігі де сондай-ақ бірдей болмай шықты. Бірінші топта (түқым қуа-лауы салыстырмалы жоғары) бір үрпақтағы сүрыптауда жыл-жу 0,6 кг (6,2 кг-нан 6,8-кг-ға дейін), ал екінші топта (тұқым қуалауы ете темен) орташа қырқылған жүн тіпті езгермеді, яғни сұрыптау тиімсіз болып шықты.
Түқым қуалауды білу — енімділікті ғылыми негізде жоға-рылату үшін де маңызды. Егер, мәселен, өнімділіктің орташа деңгейі қырқылғаи жүннен табын бойынша 1кг жоғары бола-тын қойлар мен қошқарлардың тобын сүрыптаса, онда бүл бел-гінің түқым қуалауы /г2 = 0,3 жағдайында бүл ата-аналардың үр-пақтары жүннін, өнімділігі бойынша табыиның орташа деңгейі-нен асып түседі, бірақ 1 кг емес, не бары 300 граммға асады.
lKrxO,3f 1кгхО,3\
 2
2
Түқым қуалауды білу сүрыптаудың сәйкес келетін әдістерін таңдау, сүрыптаудырі; тиімділігін анықтау, сондай-ақ өнімділік-тің артуын ғылыми негізде жоспарлау үшін өте маңызды.
34-тарау. СҰРЫПТАУ МЕТОДТАРЫ
Сұрыптау — селекцияның негізгі әдістерінің бірі болып табылады. Генетикалық әдістермен ұштаса отырып ол белгілі қасиеттері мен белгілері бар сорттармен тұқымдарды жасауға мүмкіндік береді. Мәселен, В. С Пустовойт күнбағыс тұқымы-ның майлылығын арттыру үшін 30 жыл бойына дерлік жүмыс істеп, оны 30-дан 50%-ке көбейтті. Оның шығарған сорттары дүние жүзіне танылды. Бүл табыстарға сұрыптаудың әр түрлі әдістерін қолдану негізінде қол жетті. Сұрыптау әдістерінің жү-йесіне сүрыптаудың негізгі екі типі: жаппай және жеке сүрып-тау әдістері кіреді.
1. ЖАППАЙ СҰРЫПТАУ (ФЕНОТИПІ БОЙЫНША)
Жаппай сүрыптаудың мэні. Особьтарды сыртқы белгілері (фенотипі) бойынша генотиптерін тексермей сүрыптауды жаппай сұрыптау деп атайды. Мәселен, жаппай сұрыптауда леггорн тауығы тұқымының бүкіл популяциясынан шаруашылықтарда 200—250 жүмыртқа беретін, тірі массасы 1,6 кг, түсі ақ, жұ-мыртқа басу инстинктісі байқалмаған қүстарды көбею үшін қалдырады. Бүл талаптарға сай келмейтін барлық тауықтар тобынан шығарылып тасталады. Бүл жағдайда әр тауық пен қораздың үрпағы жеке-жеке бағаланбайды, яғни бағалау тек фенотип бойынша жүргізіледі. Фенотип генотип реакциясы нор-масының көрінуі болып табылады және күшті дәрежеде сыртқы орта факторларының кездейсоқ ауытқуымен қамтамасыз етіледі, сондықтан фенотип бойынша сүрыптау генотиптерді бағалау үшін жеткіліксіз тиімді болады.
Жаппай сұрыптаудың тиімділігі. Жаппай сұрыптаудың тиім-ділігі едәуір дәрежеде белгінің түқым қуалау коэффициентіне тәуелді болады. Бүл тәуелділікті көрсету үшін мынадай мысал келтіреміз (105-сурет). Жүмыртқалағыштықтың тұқым қуалау коэффициенті /і2 = 0,25 болғанда жақсы тауықтардьщ мекиендері орта ёсеппен айына нашар тауықтардың мекиендеріне қарағанда бір жүмыртқадан сәл ғана артық жұмыртқа салып жүреді. Бі-рақ, ірі жүмыртқа салатын аналық мекиендері онан да недәуір ірі жүмыртқалар салып жүрді, а л кәлемі кіші жұмыртқа салатын аналық мекиендері одан да үсақ жұмыртқа береді. Мұның сонғысы жүмыртқа массасының түқым қуалау коэффициенті Я2 = 0,75 болуымен түсіндіріледі. Бүл жағдайда жаппай сұрып-тау бірінші ұрпақтың өзінде-ақ тиімді болады, Сонымен, жаппай сүрыптау түқым қуалауы жоғары белгілер үшін ғана тиім-ді. Жаппай сұрыптау жануарлар мен өсімдіктер популяциясын жақсартудың баяу эсер етуші құралы болып табылады, алайда ол қажетті және селекциялық жүмыстың белгілі буындарында пайдаланылады. Мысалы, ол айқас тозаңданатын өсімдіктердің селекциясында кеңінен қолданылады. Онымен халық селекция-сының сорттары жасалды. Оны қолданбаса түқымдар мен 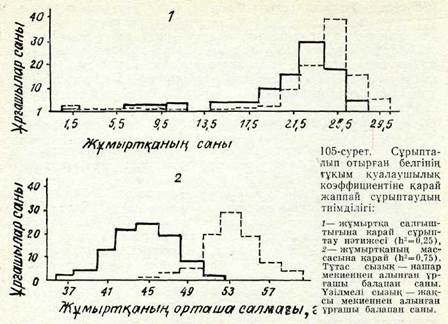
сорттар өз сапасын ауыл шаруашылық өндірісшщпроцестерінде тез жоғалтуы мүмкін.
2. ЖЕКЕ СҰРЫПТАУ (ГЕНОТИШ БОЙЫНША)
Жеке сұрыптаудың мәні. Нәтижесінде әр түрлі организмдер ұрпақтарының дербестігін жоғалтатын жаппай сұрыптаудан айырмашылығы жеке сұрыптау кезінде ең алдымен жекелеген өсімдіктін, немесе жануардың бірқатар ұрпақтар бойында ұр-пақтары бағаланады. Мұның нәтижесінде индивидуумның тұ-қым куалайтын сапаларын — ұрпақтарына қасиеттерін беру қабілетін бағалау мүмкін болады.
Жеке сұрыптау процесінде популяцияны қолдан жекелеген линияларға ыдыратады. Үрпақтарына қажетті қасиеттер бере-тін особьтарды ғана сұрыптайды, ал қалғандарын шығынға шығарады. Бүл жүмыста инбридингті жиі қолданады, ол белгі-лі бір генотиптерді сұрыптауға, сондай-ақ бағалы гендердің линияларда шоғырлануын жоғарылатуға және осының нәтижесін-де ұрпақта гомозиготалы особьтар санын көбейтуге мүмкіндік береді. Жақсы көрсеткіштері бар линияларды одан әрі селек-циялық жұмыста пайдаланады.
Жеке сүрыптаудың маңызы, генотипі бойынша бағалау, әсі-ресе бір организмнен көп мөлшерде үрпақтар алуға мүмкін-дігі бар өсімдік шаруашылығы мен мал шаруашылығының са-лаларында өте зор. Мәселен, қолдан үрықтандыруды пайдалана отырып, сперманы терең мүздатудың көмегімен бір бұқадан35000-ға дейін бұзау алуға болады. Бұл жағдайда сперманын ұрықтандыру қабілеті ұзақ жылдар бойы сақталады, яғни әл-деқашан өліп қалған аталық малдан ұрпақ алуға болады.
Соңғы жылдары көп ұрпақты тек аталықтан ғана емес, сон-дай-ақ аналықтан да, ұрықтанған аналық клетканы бір ұрға-шыдан екіншісіне ауыстырудың жетілдірілген әдісі нәтижесінде ұрпақтың үлкен мөлшерін алу мүмкіндігі туды. Бүл аса көрнек-ті асыл түқымдық малдан сақтаушы — аналыққа зиготаны ау-ыстыруға мүмкіндік береді. Бұл әдіспен бүзаулар мен қозылар алынды.
Егер қазір өте бағалы генотиптер алынатын спермалардың банкасы болатын болса, жүмыртқа клеткалар мен зиготалар-дың банкалары да пайда болатын күн алыс емес.
Жеке сүрыптауды екі әдіс арқылы жүргізуге болады.
Үрпақтары бойынша тексеру. Олардың бірі ұрпақтары бойынша тексеру болып табылады. Мысал келтірейік. Туысқан апалы-сіңлілі леггорн түқымының екі тауығы алынды. Бір.еуі (№ 9442) жылына 208 жүмыртқа, ал екіншісі (,№ 9441) 176 жү-мыртқа салды. Екі тауық та бір қоразбен шағылыстырылды. Екі тауықтың да мекиендерінің өнімділігі ескерілді, олар әр түрлі болып шықты, № 9442 тауықтың мекиендеріне қарағанда (орта есеппен 158 жұмыртқа ғана). № 9441 тауық жүмыртқала-ғыштығы ете жоғары (орташа есеппен 230 жүмыртқа) мекнен-дер берді. Егер аналықтарын сұрыптау үрпақтарының жүмырт-қалағыштығын ескермей, тек фенотип бойынша ғана жүргізіл-се, онда № 9442 тауықты қалдырған тиімді болар еді, өйткені ол көп жұмыртқа салды. Алайда үрпағы бойынша тексеріс бол-ғаннан кейін барынша бағалы генотипі бар № 9441 тауығын түқымдыққа қалдыру қажеттілігі айқын болды, өйткені ол көп жұмыртқа салу қасиетін үрпағына барынша сенімді беретін еді.
Сондай-ақ өсімдіктер селекциясында да жеке сүрыптау ке-ңінен пайдаланылады. Методикалық түрғыдан ол әсіресе өзді-гінен тозаңданатындар үшін айрықша қарапайым. Мүнда кез келген өсімдіктің түқымына оның үрпағы бойынша бағалау жүргізу үшін әр дәнді жеке сепкен жеткілікті. Мұндай сүрып-таудың мәні ол бүтіндей Иогансеннің таза линиялар туралы (25-тарауды қараңыз) іліміне негізделеді.
Түқымдық малды үрпағы бойынша бағалау мен сұрыптау — түқымдық сапаларды жетілдірудегі ең сенімді жол.
Сиб-селекция. Жеке сүрыптаудың басқа бір әдісі сиб-селекция методы болып табылады. Мүндай жағдайда сұрыптау үр-пақ бойынша емес, керісінше бүйір туысқандары — аға-інілері немесе апа-сіңлілері бойынша жүргізіледі. (Sibling ағылшын тілінде «аға-қарындас» деген үғымды білдіреді).
Сиб-селекция мысалына үялас жас қабандарды бордақылап бақылау негізінде жас мегежіндерге бордақылық сапасы бойынша сұрыптау жүргізу жатады. Бүл жағдайда жас қабандар-дың түқым қуалай анықталатын бордақылау сапасының: тезжетілгіштігі, мал азығының ақысы және т. б. көрсеткіштері бойынша жас мегежіндерді де бағалауға негіз береді. Данияда бордақылап бақылау үшін бірұяластардың әр жынысынан екі торайдан алу қабылданған. Жақсы нәтиже берген жағдайда төлдің қалған бөлігі тұқымдыққа қалдырылады, ал нашар жағ-дайда бүкіл үяластар шығарылып тасталады. Осындай әдіспен дат селекционерлеріне 10 жылда 1 кг массанын, өсіміне жұм-салатын шығын 6,48-ден 5,52 азықтық бірлікке дейін азайтудың сәті түскен. Бүтіндей алғанда бүл үлкен экономикалық тиімділік берді. Мұндай методика жартылау ддісі деген ат пен өсімдіктер селекциясында да қолданылады. Күнбағыстың май-лылығын жоғарылату үшін гүлшоғын (себет гүлін) екіге бе-лед!. Бір жартысынан тұқымының майлылығын тексереді. Егер оның майлылығының проценті жоғары болса, онда түқымның екінші жартысынан үрпақ алынады. Осы процедураны майлылығы қашан анықталғанша түқымдарды сүрыптай отырып, ұрпақтан ұрпаққа қайталай береді (106-сурет).
Сұрыптау тиімдІлігі үшін сыртқы орта жағдайларының ма-
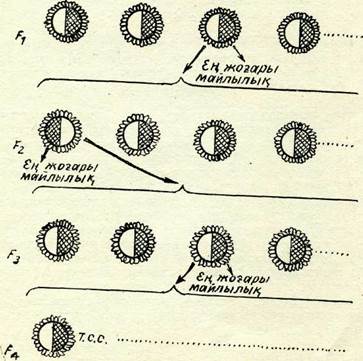
106-сурет. Сиб-селекция схемасы.
Күнбағыс кәрзеңкесінін, сызықсыз белегі майлылығы зерттелмейтін және келесі урпақ алу үшін ғана пайдаланылатын тұқымдардың жартысы; сызықты бөлегі — майлылығына қарай бағаланатын тұқымдардың жартысы маңызы.Жеке сұрыптау белгілі бір генотиптерді бағалаудың және оны жасаудың ең сенімді құралы болып табылады.
Бұл жағдайда тұқым мен сорттың олардың өздері өмір сү-ретін белгілі жағдайлар үшін жасалатынын ескеру керек. Сон-дықтан бір түқымнан немесе бір сорттан әр түрлі жағдайлар-да бірдей өнім беруді күтуге болмайды. Сұрыптау организмнің генотиггін бағалайтынына қарамастан, оның әрекеті сыртқы ортаға тәуелді болады. Сүрыпталатын организмдердің түқым қуалаушылық мүмкіндігін (генотип реакішясының нормасы) барынша айқындайтын орта жағдайларында жүзеге асырылған сүрыптау ең тиімді деп саналады. Өсімдіктердін, қуаңшылыққа төзімділігіне сүрыптауды ылғалды климат жағдайында, аязға төзімділігіне жоғары оң температура жағдайларында жүргізуге болмайды. Сыртқы ортаның сай келетін жағдайлары генотипті бағалауды женілдетеді, оны неғүрлым объективті және дәл ете-ді. Атап айтқанда, генотипті барынша толык бағалау мақсаты-мен сүрыпталатын генотиптерді неғүрлым дәл айқындауға қо-лайлы — немесе өте қолайсыз сыртқы орта жағдайларын жасау көбіне қажет болады. * * * Селекцияның генетикалық негіздерін зерттеу өсімдіктердің, жануарлар мен микроорганизмдер селекциясының эмпирика-лык тәсілдеріне ғылыми база яғни сүрыптау мен шағылыстыру-дың әр түрлі дәстүрлік әдістерінің маңызын түсіндіруге мүмкін-дік берді.
Генетика селекция үшін жаңа формалардың жасалу қарқы-нын тездететін принципті жаңа әдістерді жасады: иондайтын радиациялар мен химиялық мутагендер көмегімен түқым қуа-лайтын өзгергіштікті қолдан тудыру; гетерозис қүбылысын практикада пайдалану үшін линия аралық будандарды өсімдік-терде де, сондай-ақ жануарларда да жасау, гетерозисті бекіту әдістерін жасау; жоғары өнім беретін сорттарды жасау және әріден будандастыру кезіндегі шағылыспаушылықты жеңу үшін авто- және аллополиплоидты формалар мен синтетикалық по-пуляциялар алу әдістерін жасау; айқас тозаңданушылар (жүге-рі, сорго және басқалары) мен өздігінен тозаңданушылардың (бидай) линия аралық будан түқымдарын өндіруге жол ашқан цитоплазмалық аталық стерильділікті қолдану.
Алайда генетиканың айқын табыстарына қарамастан адам игілігіне табиғат резервтерін пайдалану үшін зерттелмеген жұмыстар әліде болса көп.
 |
I таблица
А. Пияз қабығы-нық клеткаларын-дағы митоз фаза-сы:
1 — интерфаза; 2 профаза; з— мета-фаза; 4 — анафаза; 5— телефаза.
 |
Б. Ооцит 1 трито-ныныд ядросында-ғы «лампа щет-калары» типтес хромосомалар.
 |
В. Хромосомалар-дьщ асинхронды репликациясы
 |
A. Жүгерідегі микроспорогенездің мейоздык
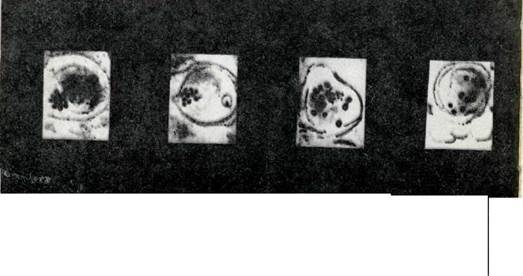
Іі. Диплоидты (1) және гаплоидты (2-4) томатта (2п-24) тозаңның пайда
болу кезіндегі I телефаза.
 |
 |
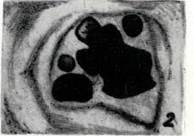
А. Мейоздьң профазасындағы бива-ленттің микрофотографиясы. Төрт хроматид көрініп тұр.
 |
 |
Б. Азиялық шегірткедегі (Locusta mirgaioria) рентгеннен кейінгі хромосома-лардың өзгерісі.
1— хромосомалар ұштарының жабысуы арқасында олардың анафазада ңалуы, жалғастырушы дәнекерлер мен фрагменттердің пайда болуы; 2— ядролардың пик-нозы — хромосомалардың жалпы массаға жабысуы.
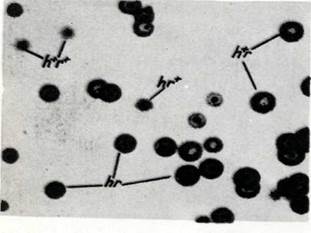 |
В. Фагтардағы рекомбинация. Фотосуретте Es-cherichia coli екі типтегі фагтарды бір мезгілде жұқтыру нәтижесінде пайда болатын негативті колониялар берілген һ+г (үлкен ақ дақтын. пайда болуын тудырады) және һг (кішкентай кара дақтьң пайда болуын тудырады). Рекомбинантты типтегі колониялар көрініп тұр: һ+г+ — кішкентай акшыл түстілер және һг — іріқара түстілер.
 |
А. Конъюгацияланатын бактериялар.
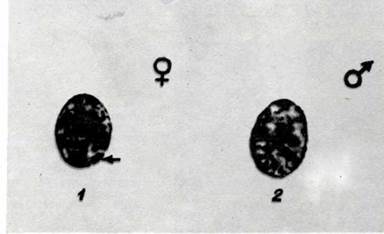
Б. Әйелдердің ұртындағы шырышты эпителий клеткасындағы (1) жы-ныс хроматині және онық еркектер клеткасында болмауы (2).
 |
 f 1<ру,
f 1<ру,
 |
 |








