 2015-06-28
2015-06-28 12943
12943Мутацияның мұндай түрі жекеленген гендерде болады және жиі кездеседі. Организмнің көптеген морфологиялық, физиологиялық және биохимиялық белгі-қасиеттері осыған байланысты өзгереді. Белгінің өзгеру бағытына қарай гендік мутацияның төрт түрін ажыратады. Олар гиперморфты-өзінің бақылауымен синтезделетін заттар мөлшерінің артуы есебінен ген қызметінің күшеюі: гиперморфты жабайы типтің аллелі арқылы бақыланатын биохимиялық өнімдер мөлшерінің азаюы есебінен ген қызметінің әлсіреуі: неоморфты-жабайы тип генінің бақылауымен синтезделетін өнімнен өзгеше заттардың тұзілуін кодтайтын мутантты аллельдің пайда болуы: аниморфты –жабайы типті аллельдің қызметіне қарама-қарсы.
Гендік мутация организм белгілерін көбінесе шамалы ғана өзгертетіндіктен оны «шағын» мутацияға жатқызады. Ол табиғи жағдайда түрлердің эволюциялық тұрғыда икемді болуына мүмкіндік туғызады және селекциялық жұмыста өсімдіктердің жаңа сортын,жануарлар тұқымдарын және микроорганизмдер штаммдарын алу үшін қажетті материал ретінде пайдаланылады.
Гендік мутацияның азотты негіздердің орын ауыстыруына және бір азотты негіздің ДНҚ құрамынан түсіп қалуына немесе үстеме келіп қосылуына байланысты болатын мутациялар негізінен екі түрін ажыратады. Мұндай өзгерістерді нүктелік мутация деп атайды.
Олар мынандай жолдармен жүреді:
1.Бір пуриннің екіншісімен немесе бір пиримидиннің басқа біреуімен ауыстырылуы.
Мұны транзиция деп атайды.

 А Г,Т Ц
А Г,Т Ц
2.Пуринді пиримидинге немесе керісінше алмастырудың. Оны трансверсия деп атайды:

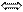
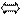
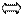 А Т, А Ц, Г Ц,Г Т
А Т, А Ц, Г Ц,Г Т
Негіздердің алмасуы үш түрлі мутанттық кодондардың түзілуіне әкеп соғады кодонның мағынасы өзгерсе (миссенс-мутация),мағынасы өзгермесе (бейтарап мутация) және мағынасыз немесе терминациялық кодондар (нонсенс мутациялар).
Берілген ген кодтайтын полипептидте миссенс мутацияның нәтижесінде бір аминқышқылы екінші аминқышқылымен ауыстырылады, бұл жерде мутацияның айқындалу қабілеті мутацияға ұшыраған доменнің функциялық маңызына байланысты. Белоктың активтік орталықтарындағы аминқышқылдарының алмасуы осы белок молекуласының түгелдей функциялық қызметінің жойылуына әкеп соғады. Адамда тұқым қуалайтын ауру орақ пішінді клеткалық анемия гемоглобулиннің B – тізбегін кодтайтын ДНҚ молекуласының гендік мутациясына байланысты. Науқастың эритроциттерінің пішіні орақ тәрізді болып,оттек тасу қабілетінен айырылады. B- тізбек 146 амин қышқылын құралған. Қалыпты және гендік мутация нәтижесінде өзгерген гендердің бір-бірінен айырмашылығы-ДНҚ-ның бір ғана негізігің өзгеруі, бұл белок молекуласында алтыншы тұрған жалғыз амин қышқылының алмасуына әкеледі: қалыпты гемоглобулиннің В тізбегінде алтыншы орында глутамин қышқылы орналасады, ал орақ пішінді клеткалы гемоглобулинде ол валин қалдығына алмасқан.
Нонсенс мутацияның нәтижесінде кез келген амин қышқылын анықтайтын кодон рибосомаларда трансляцияланбайтын немесе стоп-кодондарға айналады (UAA,UAG.UGA).Мұндай кодондардың структуралық гендердің аяқ жағында емес, іш жағында пайда болуынан белок молекуласының трансляциясының терминациясы уақытынан бұрын жүріп белок молекулаларының үзілуі болады. Нонсенс мутациясы ең көп бүлдіргіштік қабілетке ие болады, себебі өз трансляциясының терминациясы уақытынан бұрын жүрген белок молекуласы протеолиттік ферменттердің әсерінен қорғалана алмай тез деградацияға ұшырайды.
Жекленген негіздердің немесе олардың қысқа қатарларының геннің құрамында үстеме қосылуы, орын алмасуы немесе түсіп қалуы код шекарасының «оқылуының» өзгеруіне әкеп соғады.Мұндай мутациялардың табиғаты жабайы типтегі е+ генімен өзара бір-біріне супрессор фреймшифит (код шекарасы «оқылуын» өзгертетін) үш түрлі мутантты гендер кодтайтын Т4 фактың белоктарының аминқышқылдық құрамын анықтау барысында зерттелген. Кейбір жекеленген мутациялар бір уақытта көршілес бірнеше нуклеотидтер құрамындағы өзгерістерден пайда болатыны анықталды.Дәл айтқанда,код шекарасының «оқылуын» өзгертетін жекелеген мутациялар оған көрші бір емес,екі нуклеотидтердің үстеме қосылуынан пайда болатыны анықталды. Код шекарасының оқылуын өзгертетін мутациялар пайда болуы барысында оқылу бағытымен дупликация немесе делеция сайтынан кейінгі барлық триплеттер өзгеріске түседі, бұл стоп-кодондардың туындау ықтималдығын жоғарлатып трансляцияның терминациясын жүргізеді.
Хромосомалық мутациялар
Хромосомалық өзгерістер. Хромосомалардың қайта құрылуын мутацияларға жатқызу қабылданған,өйткені клеткаларда олардың болуы әдетте осы клеткалар қасиеттерінің немесе оларды пайда болатын организмдердің өзгеруімен байланысты болады.
 Хромосома ішілік өзгерістерге хромосома бөліктерінің жетіспеушілігін (дефишенсия және делекция): хромосоманың белгілі бір учаскелерінің екі еселенуін, немесе дәлірек айтқанда, скөбеюін (дупликация) хромосоманың жекелеген учаскелерінің 180 -қа аударылуы салдарынан хромосомадағы гендердің сызықтық орналасуының өзгеруін хромосома ішілік өзгерістерге хромосома бөліктерінің жетіспеушілігі ұзындықтары әртүрлі хромосома учаскелерін және оның түрлі бөліктерін қамтуы мүмкін.Егер хромосома иіндерінің бірінің соңы жойылатындай болып үзілсе, онда осы иін қысқарады.Егер онда центромералар жоқ болса, бөлініп қалған үзінді ондағы гендерімен бірге ядроның таяудағы бөлінуі кезінде жойылады. Мұндай жетіспеушілікті терминальдық немесе соңы (концевыми) деп атайды.
Хромосома ішілік өзгерістерге хромосома бөліктерінің жетіспеушілігін (дефишенсия және делекция): хромосоманың белгілі бір учаскелерінің екі еселенуін, немесе дәлірек айтқанда, скөбеюін (дупликация) хромосоманың жекелеген учаскелерінің 180 -қа аударылуы салдарынан хромосомадағы гендердің сызықтық орналасуының өзгеруін хромосома ішілік өзгерістерге хромосома бөліктерінің жетіспеушілігі ұзындықтары әртүрлі хромосома учаскелерін және оның түрлі бөліктерін қамтуы мүмкін.Егер хромосома иіндерінің бірінің соңы жойылатындай болып үзілсе, онда осы иін қысқарады.Егер онда центромералар жоқ болса, бөлініп қалған үзінді ондағы гендерімен бірге ядроның таяудағы бөлінуі кезінде жойылады. Мұндай жетіспеушілікті терминальдық немесе соңы (концевыми) деп атайды.
Кейде хромосоманың бірден екі иіні үзіледі, осының салдарынан оның екі ұшы да жойылады. Бұл жағдайда оның ашық ұштары митозда шеңбер тәрізді хромосома түзе отырып қосылуы мүмкін.
Хромосоманың бір иіні бір мезгілде екі жерден үзілгендеде хромосома бөліктерінің жетіспеушілігібайқалады. Үзілген жерлер қосылады да, хромосома қысқарады, бұл кезде ішкі учаскеде жойылады. Егер бөлініп қалған үзінді айтарлықтай үлкен болса, жойылар алдында оның ашық ұштары қосылуы мүмкін,ал метафазада ацентрлік шеңбер түзіледі.
Хромосоманың шағын учаскелерінің бөлініп қалуына байланысты микроделецияны шамалы жетіспеушілікке жатқызады. Ұсақ жетіспеушіліктер кейде фенотиптік эффект бере отырып және гендік мутацияны бейнелей отырып әдетте гомозиготалы күйде сақталады. Оларды ген мутациясынан айыратын бірден-бір көрсеткіш -оларда кері мутациялану болмайды.
Хромосома бөліктерінің үлкен жетіспеушілігі әдетте гомозиготалы күйде летальды болады,бұл хромосоманың әрбір локусының клетка мен организм тіршілігі үшін зор маңызы бар екендігін дәлелдейді. Жетіспеушіліктен гетерозиготалар ғана тіршілікке қабілетті болуы мүмкін. Бұл жағдайда жетіспеушілік арқылы анықталатын мутациялар доминанттық ретінде көрініс береді. Хромосома учаскесі жетіспеушілігінің фенотиптік эффектісі хромосоманың гентикалық системасын, гендердің орналасу реттілігін, олардың өзара байланысын бұзатындығымен түсіндіреді.
Ірі жетіспеушіліктерді генетикалық және цитологиялық әдістермен анықтауға болады.Мәселен адамның «мысықша мияулауының» синдромы сипатталған. Бұл синдром баланың сәби кезіндегі мысықтың мияулауына ұқсас ерекше жылайтынына байланысты осылайша аталған. Ақыл-ой және дене кемістігі, микроцефалия және басқа бірқатар аномалиялар осындай сипат-ты белгілер болып табылады. Цитологиялық зерттеулер арқылы 5-хромосоманың біріндегі қысөа иіннің шамалы интеркалярлық делециясын табады. Бір шеңберлі 5-хромосомасы бар ауру адамның осындай аномалиясы сипатталған. Бұл жағдайда шеңбер түзуде де 5-хромосоманың қысқа иінінде делеция байқалады. Адам, сондай-ақ жануалар мен өсімдіктер үшін де мұндай мысалдар көптеп сипатталған.
Егер қалыпты жағдайда әрбір хромосомада ген бір дозамен берілсе, онда ол екі еселенгенде немесе көбейгенде дупликацияда - геннің дозасы тиісінше 2 есе және одан да артады. Мысалы,егер қалыпты хромосомадағы гендер АВС тәртібімен орналасса,гендердің бірінің дупликациясы кезінде АВВС немесе АВВВС күйі пайда болуы мүмкін және т.с.с. Сірә, құрамында бірнеше ген болатын хромосоманың ұқсас учаскелерінің және т.б табылған. АВСАВСАВС дупликацияның неғұрлым жиі кездесетін жағдайы болса керек.
Дупликацияланған учаскелер хромосоманың көршілес учаскелерінде ғана болып қоймайды,сонымен қатар бүкіл хромосомаға таралуы, ал кей жағдайларда басқа хромосомаларға да орын ауыстыруы мүмкін. Сірә, ұқсас учаскелердің көбеюі хромосомалар мен түрлер эволюциясындағы кең таралған құбылыс.
Хромосома ішінде инверсия түзілуі үшін ол екі жерден үзілуі қажет. Инверция көбінесе рецессивті летальдв эффектімен байланысты, сондықтан олар гомозиготалы күйде сақталмайды және олар әдетте гетерезиготадан табады.
 Алайда летальды эффектімен байланысы.Егер учаскелерінің бірінде гендер АВСД тәртібімен қалыпты орналасса, олардың ВС учаскесін 180 -қа айналдыру жоқ инверсиялар да кездеседі.
Алайда летальды эффектімен байланысы.Егер учаскелерінің бірінде гендер АВСД тәртібімен қалыпты орналасса, олардың ВС учаскесін 180 -қа айналдыру жоқ инверсиялар да кездеседі.
Инверцияны алып хромосомалардың немесе мейоздың пахитена сатысында цитологиялыщ әдіспен табуға болады. Гомозиготалы инверцияда хромосомалар коньюгациясымен кроссинговер қалыпты жүзеге асады.
Геннің басқа гендер системасындағы жағдайына байланысты оның әсерінен өзгеруін А.Стертевант жағдай эффектісі деп атауды ұсынды. Бұл генді дербес бірлік ретінде емес, генотиптің бүкіл системасының, бір бөлігі ретінде қарастырудағы алғашқы талпыныс еді. Қазіргі кезде мұндай көзқарастың дұрыстығын дәлелдейтін көптеген фактілер бар.
Хромосомалық қайта құрылудың, жәрдемімен генотиптердің жаңа системалары түзіле алады. Мәселен, тіршілікке қабілетті, транслокациясы, инверциясы немесе дупликациясы бойынан гомозиготалы формалар пайда болатын жағдайларда ол белгіні бір тіршілік ету жағдайларына бейімделген болып, көбейіп, одан соң жеке түрге оқшаулануы мүмкін. Бұл жаңа түрдің бұрынғы гендері сақталады, бірақ олар не басқа тіркесу тобында болуы керек немесе олардың хромосомада орналасу реті өзгеруі керек. Демек, хромосомалық қайта құрылу эволюция үшін де маңызды рөл атқарады.
Геномдық мутациялар
Әрбір түр үшін хромосоманың саны, формасы мен мөлшері систематикалық белгілер болып табылады. Кариотиптің негізгі бірлігі хромосомалардың гаплоидты жиынтығы, яғни гомологты хромосомалардың әрбір жұбынан біреуі ғана жиынтық. Мұндай гаплоидтық жиынтыққа шоғырланған гендер тобын геном деп, ал гаплоидтық жиынтықтағы хромосомалар санын негізгі сан деп атайды да, оны n әрпімен белгілейді.
Митоз бен мейоз ұрпақтан ұрпаққа берілетін хромосомалар санының тұрақтылығын қамтамасыз ететін клетка бөлінуінің аса дәл механизмі болып табылады. Алайда кейбір жағдайларда бұл механизм бұзылады, ол хромосомалардың клетканың полюстеріне қарай әркелкі ажырауынан - хромосомалардың ажырамауынан, сондай-ақ хромосомалардың цитокинезсіз (эндомитоз) екі еселенуінен байқалады. Мұндай бұзылудың нәтижесінде хромосомалар саны өзгерген клеткалар пайда болады.
Тұтас гаплоидты жиынтық санының немесе жекелеген хромосомалардың артуы немесе кемуі есебінен хромосомалар саны өзгеруі мүмкін. Гаплойдты жиынтықтары тұтас көбейген организмдерді полиплоидтар деп атайды. Хромосомалардың саны гаплоидты жиынтыққа еселі болмайтын организмдерді анеуплоидтар немесе гетероплоидтар деп атайды.
Организм клеткаларындағы хромосомалар санының өзгеруі оның белгілері мен қасиеттерінің өзгеруімен қатар жүреді, сондықтан оларды геномдық мутациялар де атайды.
Полиплондия. Полиплондия- гаплоидты жиынтыққа қарағанда еселенген хромосомалар санына артатын геномдық мутациялар. Хромосомаларының, гаплоидты жиынтықтарының саны әр түрлі болып келетін клеткаларды: 3n - триплоидты, 4n- тетраплоидты деп және т. с.с деп атайды. Полиплоидты клеткалардан дамып жетілген организмдерді тиісінше триплоидтар,тетраплоидтар деп атайды және т.с.с.
Гомозиготалы организмнен пайда болған тетраплоид та гомозиготалы болады. Егер жиынтықтың көбеюі гомологты хромосомаларында белгілі бір гендердің әр түрлі аллельдері болатын буданды организмде жүзеге асатын болса, онда түзілетін:тетраплоид та осы гендер бойынша гетерозиготалы болады.
Соматикалық клеткалар полиплоидизацияланған жағдайда клеткалар организмнің бастапқы полиплоидтық клеткадан дамыған бөлігінде ғана полиплоидты болады да организм химер-лы болып шығады. Егер полиплоидизация зиготаның бірінші бөлінуінде жүрсе, онда ұрықтың барлық клеткалары болып шығады. Барлық хромосомалардың мейозда ажырамауы хромосомаларының саны редукцияланбаған гаметалардың түзілуіне әкеп соғады, мұндай гаметаларда хромосомалар жиынтығы бірден емес, екіден болады. Хромосомалардың жиынтығы редукцияланбаған гаметалар ұрықтануға қатысқанда полиплоидты организмдер пайда болуы мүмкін.
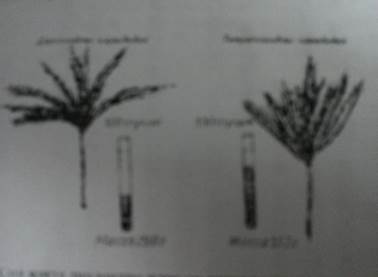
9-сурет. Сол жақтағы диплоидты және оң жақтағы тетраплоидты қара бидай.
Полиплондия. Полиплондия организм белгілерінің өзгеруіне әкеп соғады, сондықтан да ол эволюция мен селекцияда әсіресе өсімдіктер эволюциясы мен селекциясында өзгергіштіктің маңызды көзі болып табылады. В.С.Федоров селекциясындағы диплоидты және тетраплоидты қара бидай өсімдігінің, фотосуретінде бұл жақсы көрінеді (9-сурет). Тетраплоидты форманың сабағы,масағы дәуір қуаты және дәні ірі болып келеді,бұл шаруашылық тұрғыдан алғанда оны барынша тиімді етеді.
Бидай туысы (Triiiсит) бірнеше түрден тұрады, олар хромосомаларының саны бойынша сондай-ақ өсімдіктің қасиеттері мен белгілері жөнінен үш топқа бөлінеді. Бірінші топқа,мысалы, соматикалық клеткаларында саны 14-ке тең диплоидты хромосомалары бар бір дәнектілер (Т. топососсит) жатады. Екінші топқа 28 хромосомасы бар қатты бидай (Т.durumn) жатады. 42 хромосомасы бар жұмсақ бидай (aestivum) үшінші топқа кіреді. Егер бидайда хромосоманың саны n=7 болса, бір дәнектәлер диплоидты (7х2=14), қатты бидайлар тетраплоидты(7х4=28), ал жұмсақ бидайлар -гексаплоидты (7х6=42) болып шығады. Сұлы (Avena) туысы ішінде және басқа көптеген өсімдіктерде полиплоидтардың осындай қатары белгілі.
Хромосомаларының негізгі санға есептелгенхромосомаларының саны еселеніп артып отыратын қатар құрайтын туыстық түрлер тобын полиплоидтық қатар деп атайды. Полиплодтық қатар екі мүшелі және көп мүшелі болуы мүмкін.
Автополиплоидия. Бір түрдің геномдарының көбеюі негізінде пайда болған полиплоидтарды автополиплоидтар деп атйды. Егер хромосомалардың негізгі санын (геном) А әрпімен белгілесек, онда А гаплоидқа, АА- автодиплоидтқа, ААА-автотриплоидқа, АААА-автотетраплоидқа сәйкес келетін болады және т.с.с.
Автополиплоидтар табиғи жағдайларда кез-келген әдіспен көбейетін организмдерде пайда болады. Автополиплоидтар әсіресе аномикті және вегетативті жолмен көбейетін формаларда өте бағалы, өйткені олар салыстырмалы алғанда өзгеріссіз түрде ұзақ уақыт бойы сақтала және көбейе алады. Егер бастапқы формасы гетерозиготалы болса, жыныстық жолмен көбеюде автополиплоидтар тұқым қуалаушылығы алуан түрлі формалар береді.
Автополиплоидтар мейозының диплоидтар мейозынан айырмашылығы бар. Мәселен, тетраплоидта профазада биваленттер ғана емес, сонымен қатар (барлық гомологты хромосомалар өзара коньюгациялана алатындықтан) триваленттер, квадривалент-тер және униваленттер де түзіледі. Плоидтылық едәуір жоғары болғанда барлық гомологты хромосомалардың коньюгациялануы поливаленттердің немесе мультиваленттердің түзілуіне әкеп соғады.
Егер диплоидты организм хромосомаларының бір жұбы белгілі бір ген (Аа) жөнінде гомозиготалы болса, мейоз нәтижесінде гаметалардың екі сорты 1 А: 1 а пайда болады.Автотетраплоидтың, ААаа гетерозиготалы диплоидтан пайда болған редукциялық бөлінуінде гомологты хромосомалар полюстерге мынадай қатынастарда ажырауы мүмкін: 2: 2, 3: 1, 1: 3, 4: 0, 0: 4. Осы жұптың үш, бір хромосомалы және хромосомасыз гаметалары, атап айтқанда ААа және а, Ааа және А, сондай-ақ О толық бағалы болып есептелмейді. Бұл тіршілікке қабілетсіз зиготолардың пайда болуына әкеп соғады, яғни полиплоидтардың фертильділігін кемітеді.
Бірақ тіпті гетерозиготалы ААаа автотетраплоидта хромосомалар полюстерге қарай үнемі 2: 2 қатынасында тарайтын болса, тетроплоидтардағы ажырау бұл жағдайда да диплоидтардың,моногибридті ажырауынан өзгеше болады. Берілген аллель бойынша гетерозиготалы ААаа автотетраплоид гаметалардың \ АА: ААа: \аа қатынасындағы үш типін түзеді: Ғ2,дефенотип бойынша ажырау 35: 1 болып швғады, яғни осы секілді диплоидтан (3: 1) айтарлықтай өзгеше болады. 35: 1 қатынасындағы ажырау автотетраплоидты өсімдіктерге жүргізілген тәжірибелер арқылы әлденеше рет дәлелденді, атап айтқанда, ол тұңғыш рет гүлдің күрең қызыл және ақ бояуының тұқым қуалаушылығын зерттеуде сасық меңдуанамен (Датига) жасалған тәжірибелерден алынды. Диплоидтардың 1\2-і ғана гетерозиготалы болатын, болса, Ғ2-нің 36 буданының 34-і гетерозиготалы болуы айтарлықтай маңызы бар жағдай.Гексаплоидта Ғ2 ажырау 399: 1 болады.
Бұдан диплоидияға қарағанда полиплоидия гетерезиготалық-ты едәуір дәрежеде сақтайды деген қортынды шығады.
Аллополиплоидия. Әр түрлі түрлер геномдарының көбеюі нәтижесінде пайда болатын полиплоидтарды аллополиплоидтар немесе амфиплоидтар деп атайды. Мысалы,түр аралық буданда А және В геномдары табысатын болса,одан алынған аллотетраплоид ААВВ болады. Аллополиплоидияны басқаша түрде будандық полиплоидия деп атайды. Оның зор практикалық маңызы бар.
Алшақ будандар көп ретте ұрықсыз болып шығады (мысалы, қара бидайдың бидаймен, шомырдың капустамен буданы және т.б). Бұл құбылыстың себептерінің бірін қарастырайық. Шомыр мен капустаның геномдары R және В табысты делік, бұл жағдайда шомыр-капуста буданында әр түрлі екі геном- Я және В болады. Шомыр будан зиготасына 9 хромосомадан тұратын геномын, капуста да 9 хромосомадан тұратын В геномын енгізеді. Мұндай алшақ Ғ1 буданының соматикалық клеткаларындағы хромосомалардың жалпы саны 18. Мейоз профазасына жыныс клеткаларының түзілуі процесінде гомологты хромосомалар коньюгациясы жүруі тиіс.
Алайда шомыр хромосомаларының капуста хромосомалары ішінде гомологтары болмайтындықтан, мейозда олардың әрқайсысы өзін унивалент ретінде жеке ұстайды. Мейозда көрсетілген будан клеткаларында 18 унивалентті санауға болады. І анафазада олар полюстерге қарай тәртіпсіз таралады да, осының нәтижесінде хромосомаларының саны 0-ден 18-ге дейінгі түрліше шамада кездесетін гаметалар түзіледі.Олардың көпшілігі теңестірілмеген болып шығады,яғни олардағы хромосомалардың саны негізгіге қарағанда еселенбеген, сондықтан тіршілікке қабілетсіз болып шығады.
Алайда мұндай будандағы аналық, сондай-ақ аталық гаметалардың кейбір бөлігінде 9В – 18 хромосома болады. Бұл гаметаларды редукцияланбаған деп атайды. Редукцияланбаған гаметалардың ұрықтану процесінде бірігуінен екі түрдің де хромосомалар жиынтығы екі еселенген зигота аллотетраплоид немесе амфидиплоид пайда болады. Онда шомыр хромосомаларының екі жиынтығы (9R+9В) және капуста хромосомаларының екі жиынтығы (9 R+9В),яғни барлығы 36 хромосома болады және оның мейозында әрбір хромосоманың коньюгацияланатын серігі болатындықтан фертильді болып шығады.
Амфидиплоидтарды алу будандастыру жолымен және будандардың хромосомалар санын екі еселендіру жолымен жаңа константтық формалар синтездеу мүмкіндігін ашты,өйткені олар бастапқы формаларды бермиді және өздеріндегі костант белгілірге аралық тұқым қуалаушылық тән болады. 20-жылдардың басында Г.Д.Карпеченко шомырды (Raphanus sativus) капустамен (Brassica oleracea) будандастыру арқылы алғаш рет туыс аралық өсімтал будан алады.Бұл өте мықты будан болып шықты,әрі ол шомыр мен капустаның белгілерін біріктірді.Ол келесі ұрпақтарда да өсімтал әрі константты күйінде қалды. Екі туыс геномдарының үйлесуі негізінде синтезделген бұл жаңа форма рафанобрассика (Raphanobrassica) немесе шомыр-капуста буданы деп аталды. 10-суретте рафанобрассиканың (4) жемісі мен хромосомалар жиынтығы келтірілген. Бұл форманың бұршаққынының жоғары бөлігі шомырдан (10- сурет,1),ал оның табаны капустадан (10-сурет,2) құрастырылған болып шықты. Бұл буданның диплоидты формасы ұрықсыз болады (10-сурет,3).
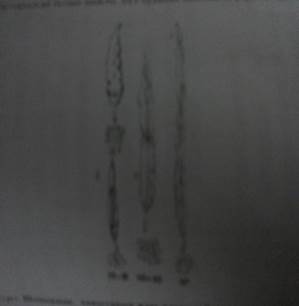
10-сурет.Шомырдың,капустаның және олардың будандарының жемістері мен хромосома жиынтықтары.
Әр түрлі геномдарды үйлестіру және полиплоидизация жолымен табиғатта жоқ жаңа формаларды синтездеуге болатыны қарастырылған мысалдан көрінеді. Мұндай жаңа формаларды шығаруды түрлердің синтезі деп атайды. Бірқатар ұрпақтар бойы сұрыптаудан кейін олар толығымен константты бола алады және оларды дербес таксономиялық бірліктер деп санауға болады.
Жануарлар полиплоидиясы. Дара жынысты жануарларда полиплоидия өте сирек кездесетіні анықталды.
Қазіргі кезде тұт көбелегінің,тритонның,аксолотльдің автополиплоидты особьтары алынды. Жібек құрты (Вомвук mori) автотетраплоидтарының ұрғашылары есімтал,ал еркетері стерильді келеді. Мұның себебі,еркектерінде (гомогаметалы жыныс) мейоз профазасында поливаленттер түзіледі, осының нәтижесінде хромосомаларының саны анеуплоидты болып келетін тіршілікке қабілетсіз гаметалар пайда болады. Ұрғашыларының (гетерогаметалы жыныс) хромосомалар коньюгациясы шектелген,поливаленттер түзілмейді,ал пайда болушы гаметалар тіршілікке қабілетті келеді. Дара жынысты жануарларда полиплоидтардың серек кездесу себептерінің бірі --олардың ұрықсыздығы.
Жыныстық көбеюі партеногенезбен алмастырылған жануарларда полиплоидия түгелдей дерлік өсімдіктердегідей жүзеге асуы мүмкін.
Жануарлардағы полиплоидтық қатар мысалына олардың аскаридадағы,жер құртындағы,қос мекенділердегі,көбелектердегі және басқалардағы көрінісін жатқызуға болады.
Жануарлар аллополиплоидтарын жасанды жолмен қолдан алу соңғы уақытқа дейін шешілмейтін проблема болып есептеліп келеді. Алайда таяуда қызметкерлерімен бірге Б.Л.Астаурров Вамвух morix x B.mandarina жібек көбелегінің түр арлық буданынан алғашқы аллополиплоид алды. Аллотетраплоидты синтездеуде жасанды партеногез әдісі пайдаланылды. Шағылыстыру процесінде екі жыныстың да әр түрінің хромосомалар жиынтығы екі еселенген формалары сұрыпталып алынды:2n B. mori + 2n B. mancdarina-аллотетраплоидтар немесе амфидиплоидтар.
Жануарлар дүниесіндегі полиполидия бірқатар тканьдердің соматикалық клеткаларында, яғни митоз жолымен мейозсыз көбейетін жағдайларда айтарлықтай кең таралған.
Қазіргі кезде генетиктердің көпшілігі жануарлар эволюциясында полиплоидия емес, хромосома арлық және хромосома ішілік қайта құрылулар негізгі рөл атқарды деген көзқарасты жақтайды.
Анеуплоидия. Анеуплоидия (немесе гетероплоидия) дегеніміз гаплоидты саны еселенбеген, хромосомалар санының өзгеруінен тұратын геномдық мутация.
Бұл құбылысты дрозофиланың жыныспен тіркескен белгілерінің, тұқым қуалауын зерттеуде К.Бриджес алғаш рет таза генетикалық әдіспен анықтады.
Жекелеген хромосомаларының саны өзгерген клеткалардың пайда болуын клетка бөлінуде хромосомалардың кейбір жұптарының тарауы бұзылатындығымен түсіндіруге болады. Мұндай бұзылушылықтар соматикалық және жыныс клеткаларында кездесуі мүмкін. Жоғарыда сөз болған, кейбір полиплоидтар мен алшақ будандар секілді гомологты хромосомаларының коньюгациясы бұзылған формаларда тұрақты анеуплоидты гаметалар пайда болады.
Егер қосымша хромосомасы (n+1) бар гамета қалыпты, гаплоидты (n) гаметамен үйлессе, зиготада бір хромосома артық болып шығады: ондағы хромосомалар саны 2 n+1-ге тең болады. Бір хромосомасынан айырылған гамета қалыпты, яғни гаплоидты гаметамен үйлессе, диплоид саны толық емес бір хромосома жетіспейтін (2 n-1) зигота пайда болады.
Хромосомаларының жиынтығы 2n +1 болатын организмді трисомик деп, 2 n-1 болатын организмдерді- моносомик деп, ал 2 n-2 болатын организмдерді - нулисомик деп атайды. Кейбір сирек жағдайларда хромосомасының белгілі бір жұбының қосымша бір хромосомасы емес,екі хромосомасы (2 n+2)-тетросомигі, үш хромосомасы (2 n+3) –пентпсомигі болуы мүмкін және т.с.с.
Мұндай санының кемуі немесе артуы хромосомалардың кез-келген жұбынан байқалуы мүмкін, сондықтан гомологті емес хромосомалардың бірнеше жұбында бір мезгілдік анеуплоидия болуы мүмкін.
Анеуплоидия белгілердің тұқым қуалау сипатының өзгеруіне ғана әкеп соқпайды, сондай-ақ ол фенотипте де белгілі бір өзгеріс туғызады. Мәселен, адамда жиынтықтың барлық хромосомалары бойынша трисомия сипатталған. Трисомия кейде толық болады, яғни бір нөмірдің үш хромосомасы қайталанады. Трисомияның мұндай жағдайы әсіресе геномның ірі хромосомаларында жиі кездеседі. Бұл жекелеген хромосомалардың генетикалық құндылығы әркелкілігін білдіреді. Ішінара трисомия негізінен алғанда инверсияның немесе геномдағы дупликацияның болуы есебінен түзіледі. Әрбір хромосоманың трисомиясы фенотиптік тұрғыдан алғанда симптомдардың белгілі
17 Дәріс
Дәрістің тақырыбы: Тұқымқуалаушылық өзгергіштікті туғызатын факторлар.
Жоспар:
1.Табиғи (секірмелі) мутагенез
2.Жасанды (индукциялы) мутагенез
3.Радиоактивті сәулелердің мутагендік әсерлері. Модификация және реакция нормасы.
4.Химиялық қосылыстардың мутагендік әсерлері.
5.Тұқым қуалайтын өзгергіштіктегі гомологиялық қатарлар заңы.
6.Модификацияның тудыратын факторларға байланыстылығын сипаттау. 7.Модификацияның бейімдеушілігі.
Дәрістің мақсаты: Тұқымқуалаушылық өзгергіштікті туғызатын факторлармен танысу.
Табиғи (секірмелі) мутагенез
Секірмелі (спонтанды) түрде болатын мутациялық өзгергіштікке белгілі бір факторлармен арнайы әсер ету арқылы емес табиғи жағдайда өздігінен пайда болатын мутациялар жатады. Де-Фриздің мутациялық теориясының қалыптасуы әртүрлі өсімдіктер мен жануарларда болатын тұқым қуалайтын өзгергіштіктерді тауып зерттеуге мүмкіндік туғызады. Жалпы мутацияның табиғатта кең таралатындығы ертеден белгілі. Мысалы, ХІХ ғасырдың бас кезінде Францияда Версаль маңында кәдімгі сарыағаштың (барбарис) әсінділерінің арасынан қызыл жапырақты формасы табылған. Кейін одан алынған ұрпақтар да қызыл жапырақты болып шыққан. Сол сияқты сасық меңдуананың тұқым қорабының тікенектері жоқ түрі табылған және ол қасиеттің ұрпақтан-ұрпаққа, тұрақты түрде беріліп отыратындығы анықталған. Спорта деп аталатын бүршікті мутацияларға кезінде Ч.Дарвин көңіл аударған болатын. Ол оның кейбіреулерін сипаттады. Мысалы, қарлыған өсімдігінің бір бұтағының өзінде бірнеше түсті жидектердің болатындығын және бадам ағашының шабдалыға ұқсас жеміс беретіндігін байқады. 1924-1925 жылдары неміс генетигі Э.Баур намазшам гүлдің түсі мен құрылысы бойынша ерекше көзге түсетін мутацияларды анықтады. Ол осы түрге жататын әрбір 1000 өсімдікке кемінде екі мутациядан келетіндігі тапты. Жануарларда да көптеген мутациялар анықталды. Олардың көпшілігі Ч.Дарвин атап көрсеткен болатын. Солардың біріне 1791ж. АҚШ-тың Массачусетс штатындағы Анкон формасында, кейіннен анкон тұқымының қалыптасуына бастау болған қысқа аяқты қойлардың шығуы жатады. Мутациялық өзгергіштік дрозофила шыбынында біршама кеңірек зерттелген. Олар денесінің түсі, қанаты, көзі, аяғының пішіндері, жыныстық белгілері, т.б. белгілеріне қатысты. Табиғи жағдайда салыстырмалы түрде алғанда мутация сирек кездеседі. Мысалы дрозофилада ақ көзді мутацияның жиілігі әр 100000 гаметаға біреуден-ақ келеді. Бір генге шаққандағы мутацияның орташа жиілігі бактерияда шамамен 1:10000000. Адамда гендердің мутацияға ұшырау жиілігі 1;200000. Әрбір жеке генге шағып қарастырғанда бұл көрсіткіштер аса жоғары емес. Бірақ жоғары сатыдағы организмдердің гаплоидты хромасома жиынтығында бірнеше мың геннің болатындығын және оның әрқайсында мутация жиілігі 1 1000 000 болып келетіндігін ескерсек мутациялы гаметаның саны оншалықты аз да болып шықпайды. Бір организмнің әртүрлі гендері түрліше жиілікте өзгере алады. Немесе бір гендер басқаларына қарағанда бірнеше рет мутациялануы мүмкін. Мұндай заңдылық жүгері эндоспермінің белгілерін анықтайтын жеті түрлі геннің секірмелі мутацияларынан аңғарылады. Бұл өсімдіктің боялған алейрон қабатының гені бойынша 500 гаметаға шаққанда бір мутация пайда болатын болса, ал қатпарлы эндоспермнің генін алсақ бір миллион гаметаға бір ғана мутациядан келеді. Секірмелі мутацияның мөлшері организмнің физиологиялық жағдайына және клеткада болатын биохимиялық өзгерістерге де байланысты. Мысалы, М.С.Навашин мен Г.Штуббеннің айтуы бойынша тұқымды қамбада ұзақ уақыт бойы сақтау нәтижесінде олардың ескіруі себепті мутациялық өзгергіштің пайда болу жиілігі артады. Секірмелі мутацияның пайда болу себебінің біріне, клеткада кейбір заттардың биосинтезіне кедергі келтіретін мутагендік қасиеті бар бөгде қосылыстардың жинақталуына да жатқызуға болады.
Жасанды (индукциялы мутагенез) Жасанды немесе индукциялы мутация деп белгілі бір факторлармен арнайы әсер ету арқылы пайда болатын тұқым қуалайтын өзгергіштікті айтады. Ондай мутагендік факторларға радиоактивті сәулелер, температура, ультра күлгін және лазер сәулелері, сол сияқты кейбір химиялық қосылыстар (колхицин, этиленимин, азотты спирт, акридиндер, метилметансульфанат т.б.) жатады. Өсімдіктер, жануарлар және микроорганизмдермен жүгізілген көптеген зерттеулердің нәтижесінде осы аталған факторлармен әсер ету арқылы мутация тудыруға болатындығы анықталды. Мутациялық өзгергіштікті қолдау жасауда әсіресе, радиоактивті сәулелердің мутагендік қасиетін зерттеп білуджің ерекше маңызы бар. Радиоактивті сәулелердің мутагендік әсерлері Тұқым қуалаушылық қасиеті радиацияның әсерімен өзгертуге болатындығын тұңғыш рет 1925 ж. Г.А.Надсон мен Г.С.Филиппов дәлелдеді. Олар радий сәулелерімен әсер ету арқылы саңырауқұлақтардың жаңа мутантты формаларын алды. Одан кейін 1927 ж. Г.Меллер дрозофила шыбынынан, ал 1928 ж. Л.Стадлер жүгері мен арпа өсімдігінен рентген сәулелерінің мутагендік әсерін байқады. Радиоактивті сәулелердің мутагендік қасиетін түсіну үшін ең алдымен радиацияның әсерінен клетка ядросындағы соның ішінде хромосомалардағы болатын өзгерістерді білу керек. Клеткада жүретін тұқым қуалау процесі құрамына негізінен дезоксирибонуклеин қышқылы енетін хромосомалармен тығыз байланысты. Олай болса радиацияның әсерінен ең алдымен ядролық құрылымдар өзгереді. Соның салдарынан барып мутация пайда болады. Радиоактивті сәулелердің төменгі дозалары нүктелік мутация тудырады, яғни ол жағдайда хромосоманың шағын ғана бөлігінде немесе жеке бір генде өзгеріс болады. Ал жоғары дозаларда хромосомалардың үзілуі, орын алмастыруы сияқты күрделі өзгерістер пайда болады. Мұндай жағдайда жаппай мутациялану жүріп зат алмасу процесі өзгереді, бара-бара клеткалардың тіршілігі жойылып ақырында организм өлімге душар болады. Радиоактивті сәулелердің әсерінен пайда болатын мутациялар организм үшін көбінесе зиянды болып келеді. Мысалы, адамда тұқым қуалайтын психикалық өзгерістерді, қан ауруларын, бедеулік т.б. тудырады. Қазіргі кезде радиацияның тұқым қуалаушылыққа әсер етуінің механизмдерді біршама зерттелген деуге болады. Соның бірі организмдердің радиацияға сезімталдығы. Мысалы, дрозофила шыбындарының радиацияға сезімталдығы тышқандарға қарағанда 10-20 есе кем болады. Себебі, ересек шыбындарда клеткалардың бөліну қарқыны тышқандар мен басқа сүтқоректілерге қарағанда көп болып келеді. Ал радиацияға сезімталдық клетканың бөліну қабілетіне тікелей байланысты нәрсе. Радиауияға әсіресе ит пен маймыл өте сезімтал болып келеді. Мұндай зерттеулер адамның сәулеге сезімталдығы туралы болжам жасауға негіз бола алады. Сәуленің әсерінен болатын генетикалық зақымдалу организмнің қандай түрге жататындығына да байланысты болады. Генетикалық радиосезімталдықтың түрге тән ерекшеліктері тәжірибе жүзінде зерттелген. Мысалы, теңіз шошқасының жыныс бездерінде 4р (рентген) дозаның өзі-ақ. Жыныс клеткалары ядроларының өзгеруіне әкеп соққан, ал мұндай дозада тышқандар мен үй қояндарының жыныс клеткаларынан ешбір өзгеріс байқалмаған. Маймылдарда 100-200р. дозамен әсер еткен кезде жыныс клеткаларының ұрықтану қабілеті жойылады, ал тышқандарда мұндай жағдай 400р. дозада ғана байқалады. Адам баласына мутацияның жартылай доминанттылығы тән болып келеді. Ол әрине, болашақ ұрпақ үшін қауіпті, себебі генетикалық кемістіктердің жүзеге асуын тездетеді. Осыған байланысты сәулеленген адамдардың бірінші ұрпағының өзінен тұқым қуалайтын аурулар байқалады. Бұған Нагасакидегі атом жарылысынан зардап шеккендер дәлел бола алады. Сонда радиацияның әсеріне ұшыраған ата-аналардың балаларының 80 пайызынан әртүрлі тұқым қуалайтын ауру мен кемістіктер байқалған. Радиацияның әсерінен дене (сомалық) клеткаларында қатерлі ісіктің (рак) пайда болуы да негізінен олардағы генетикалық құрылымдардың өзгеруіне байланысты. Рак радиацияның әсерінен бірден болмайды, өрганизм сәулеленген соң біраз уақыттан кейін барып байқалады, себебі оның алдында күрделі өзгерістер жүреді. Өсімдіктер мен жануарлардың дене клеткаларында болатын мутацияларды арнайы цитогенетикалық әдістермен зерттейді. Өсімдіктердің тұқымдарына, тозаңдарына немесе вегетативтік мүшелеріме радиациямен әсер ету арқылы әртүрлі мутанттарды алуға болад. Олар түрлі морфологиялық, физиологиялық немесе биохимиялық қасиеттерінің өзгеретіндігімен сипатталады. Қазіргі кезде өсімдіктердің 130-дай түрінен 31000 мутация анықталған, соның ішінде бидайдан 11000 мутация, күріштен – 3400, арпадан – 4500, ас бұршақтан – 3200. Мәдени өсімдіктерден алынған мутантты формалардың көпшілігі шаруашылық жағынан пайдалы болып есептеледі. Мысалы, біз радиоактивті кобальттың (Со60) гамма сәулелерімен әсер ету арқылы жаздық бидайдың Қазақстан-3, Қазақстан-126 сорттарының қысқа сабақты болғандықтан жатып қалмайды, тік өсіп тұрады. Осындай жолдармен мақтаның, қарабидайдың, арпаның, томаттың т.б. дақылдардың ауруға, қуаңшылыққа төзімді, мөл өнім беретін көптеген мутантты сорттары алынған. Химиялық қосылыстардың мутагендік әсерлері Химиялық заттардың әсерінен тұқым қуалайтын өзгергіштіктің пайда болатындығы радиоактивті сәулелердің мутагендік қасиеттері ашылғаннан кейін көп кешікпей белгілі болды. 1932ж. В.В.Сахаров дрозофила шыбынының жұмыртқаларын иодты калийдің 10 процент ерітіндісімен өңдегенде оның мутация тудыратындығын анықтады. 1938ж. М.Е.Лобашев сол объектіден аммиактың мутагендік әсерін байқады. Кейінірек мутация тудыратын қасиеті бар, бірқатар химиялық қосылыстар табылады. 1946 ж.Ш.Аэрбах пен Д.Робсон қыша газының (ипирт) мутагендік әсерін анықтады, ал И.А.Раппопорт, формальдегид пен қарапайым гетероциклді қосылыс – этилениминнің мутагендік қасиетін тапты. Қазіргі кезде 500-ден астам әртүрлі мутагендік қосылыстардың бар екендігі белгілі. Олардың қатарына диметилсульфат, диэтилсульфат, нитрозоэтилмочевина, гидроксиламин, азотты қышқыл, бромурацил, акридиндер т.б. жатады. Химиялық мутагендер көбінесе гендік мутациялар тудырады. Бұл жағдайда ДНК репликациясы бұзылады, кейде оның жіпшелері үзіліп нәтижесінде күрделі хромосомалық өзгерістер пайда болды. Радиациялық мутагенез сияқты химиялық мутагенез де өсімдік селекциясында пайдалы мутанттар алу үшін кеңінен колданылады. Тұқым қуалайтын өзгергіштіктегі гомологиялық қатарлар заңы. Толып жатқан тұқым қуалайтын өзгерістердің арасынан систематикалық тұрғыда бір-біріне жақын тұрған түрлерде кездесетін мутациялардың ұқсас болып келетіндігі анықталды. Атақты генетик Н.В.Вавилов осы мәселені жан-жақты зерттеп, соның нәтижесінде өзінің «тұқым қуалайтын өзгергіштіктегі гомологиялық қатарлар» деп аталатын заңын қалыптастырды. Бұл заң бойынша шығу тегі жағынан бір-біріне жақын систематикалық топқа жататын, морфологиялық және физиологиялық қасиеттері жағынан да ұқсас болып келеді. Мысалы, әртүрлі астық тұқымдастар – күріш, бидай, арпа, сұлы, тары, жүгері, бидайдықтар тұқымның түсі мен пішіні, масағындағы қылтанағы, ерте пісетіндігі, суыққа төзімділігі, сабағының ұзындығы т.б. қасиеттері жөнінен тұқым қуалайтын өзгергіштіктің ұқсас (гомологиялық) қатарлары болатындығы анықталды. Сонда бір түрде болатын мутациялық өзгергіштіктің болатындығын алдын-ала болжауға болады. Бұл заң мынандай формула арқылы көрсетіледі. І. 4(а+в+с...) І.2(а+в+с...) І.3(а+в+с...) І әрпімен жақын туыстар мен түрлер, ал жақшаның ішіндегі әріптермен тұқым қуалайтын белгілердің ұқсас қатарлары белгіленеді. Гомологиялық өзгергіштік болуының негізінде екі себеп жатыр: 1. Жақын түрлер мен туыстардың генетикалық құрылымның ұқсастығы мен олардың шығу тегінің бар екендігі; 2. Белгілі бір сыртқы орта жағдайларына жүргізілетін сұрыптаулардың әсері. Тұқым қуалайтын өзгергіштіктің гомологиялық қатарлар заңы селекцияда кеңінен қолдалынады. Ол көптеген тұқым қуалайтын өзгергіштіктердің ішінен қажеттілерін дұрыстаудан алуға, бір түрде дәл сондай мутацияны қолдан жасауға мүмкіндік береді. Мысалы, ХХ ғасырдың 20-жылдарына дейін қатты бидайдың тек қылтанақты түрлерінің ғана бар екендігі белгілі болып келген. Ал жұмсақ бидайдың да сондай қылтанақсыз түрлерінің болатындығы қатты бидайдың да сондай қылтанақсыз түрлерін алуға мүмкіндік туғызады. И.В.Вавилов Абиссинаға барған сапарында қатты бидайдың сондай түрлерін тапты, ал белгілі селекционер А.П.Шехурдинкатты бидайдың қылтанақты сортын қылтанақсыз жұмсақ бидаймен будандастыру жолымен қатты жаздық бидайдың қылтанақсыз сортын шығарды. Тұқым қуалайтын өзгергіштіктің гомологиялық қатарлары жануарларда да кездеседі. Альбиностар (түстің ақ болуы) теңіз шошқасында, қоянда т.б. кемірушілердің барлығында болады. Модификациялық өзгергіштік Қандай болмасын генотипі жағынан бірдей, бірақ әртүрлі сыртқы орта жағдайларында өсіп-дамыған организмдердің фенотиптері түрліше болады. Особьтардың осылайша фенотип жағынан әртүрлі болып өзгеруін модификациялық өзгергіштік деп атайды. Модификациялық өзгергіштіктің өзі белгілі бір шамада өзгере алу қабілеті. Мұндай қабілет генотип арқылы анықталады. Реакция нормасын зерттеу үшін генетикалық жағынан біртекті материал алып, оны әртүрлі ортада өсіру керек. Бұл мақсатқа өздігінен тозаңданатын өсімдіктің таза сорты немесе адам мен жануарда болатын бір жұмыртқадан дамыған егіздер алынады. Модификациялық өзгергіштікке мысал ретінде жебе жапырақты келтіруге болады. Оның судың бетіндегі жапырақтары жебе тәрізді, суда қалқып жүретіндері жүрек тәрізді, ал судың түбінде болатын жапырақтары ғана таспа тәрізді. Жебе жапырақтың генотипі сыртқы орта жағдайларының әсеріне байланысты белгілі бір мөлшерде жапырақ пішінін өзгерте алалды. Модификациялық өзгергіштіктің адаптивтік мәні бар. Бүағн мысал ретінде дрожжа (ашытқы) клеткаларының әдеттегіден өзгеше субстрат галоктозаны ашыту қабілетіне ие болатындығын алуға болады. Әдетте ашытқылар глюкозаны ашытады. Егер оларды тек галоктаза ғана бар ортаға апарса, оны ашыта бастайды. Бұл жерде басқа ортада, яғни галоктаза жағдайында ашытқы клеткалары осы аталған субстратты ашытуға қатысатын арнаулы фермент синтездейді деп тұжырымдауға болады. Егер осы жаңа ортаға адаптацияланған клеткаларды қайтадан апарса, оны ашыту қабілеті қайтадан қалпына келеді. Организм белгі мен қасиеті даму қабілетін де тұқым қуалап алады, ал оның қалыптасуы генотип пен сыртқы ортаның өзара әрекеттесуіне байланысты. Мысалы, бір тұқымға жататын, бірақ әртүрлі шаруашылықта өсірілген сиырлар түрліше мөлшерде сүт беруі мүмкін. Біреуінде жылына 2105-тен 3203кг болса, екіншісінде 3003-5000 кг сүт береді. Бұл олардың генотипі және сыртқы орта жағдайларына яғни азығына, күтіміне байланысты. Ал сол бір тұқымға жататын сиырлардың әртүрлі орта жағдайларында берген сүтінің мөлшеріндегі айырмашылық (2105-5000кг) олардың ата-анасынан тұқым қуалап алған реакция нормасын көрсетеді. Әртүрлі белгілерде реакция нормасының мөлшері түрліше болады. Мысалы, адамның кейбір белгілерінің, атап айтқанда қан тобының, шаш түсінің реакция нормасы төмен де, ал бойы мен салмағының т.б. белгілерінің реакция нормасы жоғары болып келер тұқымға жататын, бірақ әртүрлі шаруашылықта өсірілген сиырлар түрліше мөлшерде сүт беруі мүмкін. Біреуінде жылына 2105-3203 кг болса, екіншісінде 3003-5000 кг сүт береді. Бұл олардың генотипі және сыртқы орта жағдайларына яғни азығына, күтіміне байланысты. Ал сол бір тұқымға жататын сиырлардың әртүрлі орта жағдайларында берген сүтінің мөлшеріндегі айырмашылық (2105-5000) олардың ата-анасынан тұқым қуалап алған реакция нормасын көрсетеді. Әртүрлі белгілерде реакция нормасының мөлшері түрліше болады. Мысалы, адамның кейбір белгілерінің, атап айтқанда қан тобының, шаш түсінің реакция нормасы төмен де, ал бойы мен салмағының т.б. белгілерінің реакция нормасы жоғары болып келеді.
Бақылау сұрақтары:
1.Табиғи жасанды мутациялар қалай пайда болады?
2.Ч.Дарвиннің зерттеген қандай мутациялары табиғи мутацияларға жатады?
3.Т.Морган зерттеген қандай мутациялар табиғи мутациялар мысалына жатады?
4.Жасанды мутациялар не тудырады?
5.Химиялық мутагендік заттарды бірінші кім ашқан?
6.Тұқым қуалайтын өзгергіштіктегі гомологиялық қатарлар заңы қалай тұжырымдалады?
7.Тұқым қуалайтын өзгергіштіктегі гомологиялық қатарлар заңын ашқан орыс ғалымы?
Пайдаланған әдебиеттер:
1.Ингс-Вечтомов. С.Г.Генетика сосновами селекции, М.Высшая школа,1989.
2.Лобашев М.Е.Ватти, К.Е.Тихомирова, М.М.Генетика с основами селекция. М. Просвещение,1979
3.Ватти К.А.Тихомирова, М.М.Руководства к практическим занятиям по генетике. М.Просвещение.1979,1972
4.Лобашев М.Е.Генетика. Изд-во ЛГУ. Генетические 1969
5.Медведев Н.Н.Практическая генетика. М.Наука 1966
6.Мұхамбетжанов К.К., Далабаев Б.А, Өтешева Г.А. Генетикадан практикалық сабақтар. Алматы Ғылым 2004
7.Мұхамбетжанов К.Қ. Генетика Алматы 2005.
18 Дәріс
Дәрістің тақырыбы: Онтогенездің генетикалық негіздері.
Жоспар:
1. Дифференцировканың (жіктелудің) генетикалық негіздері
2. Алғашқы жіктелу (дифференцировка)
3. Геннің әрекеті
4. Онтогенетикалық адаптация
5. Онтогенездің дискреттілігі (оқшаулығы)
6. Жіктелудің генетикалық механизмдері
Дәрістің мақсаты: Организмнің онтогенетикалық дамуының генетикалық заңдылықтарын оқып үйрену.
Дифференцировканың (жіктелудің) генетикалық негіздері
Көп клеткалы организмнің жеке дамуының және өсуінің негізіне клеткалардың митоздық бөлінуі жатады. Организмнің әртүрлі ұлпаларының, атап айтқанда, бұлшық еттің, терінің, т.б. органдар клеткаларының бөліну жолдары бірдей, яғни митоздық жолмен бөлінеді. Соған байланысты олардың генотиптері де ұқсас болуы керек. Ал бұлай болған жағдайда онтогенез барысында ұлпалар мен клеткалар жіктелуінің генетикалық механизмі қандай деген сұрақ туады. Мұндай күрделі сұраққа жауап бере алатын тек онтогенетика. Онтогенезді генетикалық тұрғыдан зерттеу белгінің қалыптасуына ген инвени әсерге талдау жасаудан басталады. Ол бір ген – бір белгі деген принципте жүргізіледі. Бұл мәселені соңғы деректер тұрғысынан былайша жазуға болады; ген (ДНК)-РНК-белок- белгі. Өсімдіктер мен жануардар онтогенезі кезінде өсу, ұлпалардан жіктелу және морфогенез, яғни органдардың қалыптасуы сияқты процестер жүреді. Алғашқы жіктелу (дифференцировка) Жануарда алғашқы морфологиялық жіктелудің жұмыртқа клеткасының цитоплазмасы мен оның үстіңгі қабаты (кортекстің) құрылымына байланысты болатындығы белгілі. Мұның дәлелі ретінде мысалы кейбәр омыртқасыздар мен амфибиялардың ядросыз жұмыртқаларының өзі бластула стадиясына дейін даму қабілетін сақтай алады. Кортекстің өзі дискретті (оқшау) түрде функция атқара алады. Оның эктодерма, мезодерма және гаструляция процесі басталатын зоналары болады. Сонымен жұмыртқа клеткасы ұрықтанғанға дейін де жіктеледі. Жұмыртқа ұрықтанғаннан кейін, оданәрі тағы да жіктеледі, соның негізінде ұрық дамиды. Сөйтіп, жұмыртқа клеткасының қалыптасуына аналық диплоидты организмдегі барлық гендердің жиынтығы қатысады. Ал мейоздық бөлінуден кейін онда хромосомалардың гаплоидты жиынтығы ғана сақталады. Сонымен қатар диплоидты аналық организмде түзілген гендік өнімдер цитоплазмада да болады және олар жұмыртқа клеткасының дамуының алғашқы фазасын қамтамасыз етеді.Себебі цитоплазмада аналық организм алдын-ала әзірленген, ұрықтың дамуына қажетті белоктың синтезіне матрица болып есептелетін ДНК мен информациялық РНК болады. Сөйтіп клетканың кейбір цитоплазмалық органоидтардың өзінде тұқым қуалайтын информацияны алып жүре алатын, ядролық нуклеин қышқылдары сияқты, ДНК мен РНК кездеседі. Нағыз онтогенез ұрықтанудан басталады. Аталықорганизмнен келген гендер де сол ген бастап қызмет атқара бастайды. Егер гендер бүкіл онтогенезді, яғни организмнің барлық белгілері мен қасиеттерінің қалыптасуынан бақылап отырады десек, «дамудың әртүрлі кезеңдерінде бір мезгілде бүкіл гендер, яки болмаса олардың кейбіреулері ғана қызмет атқара ма және ол немен аяқталады?» - деген сұрақтар туады. Бұл сұрақтарға жауап ретінде мынандай тәжірибелердің нәтижесін келтіруге болады. Бақаның жұмыртқа клеткасының ядросын алып тастап, оның орнына микропипетканың көмегімен дамудың соңғы саьысындағы (бластула, гаструла сияқты) ұрық клеткасының ядросы салынған жағдайда егер донор клеткасының яросы жіктелу процесін өткерген болса, онда оны алмастырып салғанда реципиент – жұмыртқадан (қабылдап алушы) қалыпты ұрық дамымайды. Ал егер донор жіктелмеген болса, онда реципиент – жұмыртқа клеткасынан қалыпты ұрық дами алмайды. Бұдан гаструляция кезінде ядроның жіктелетіндігін және оның қайтымсыз процесс екендігін көреміз. Мұнан шығатын қорытынды дамудың әр кезеңінде белгілі бір арнаулы гендер ғана қызмет атқара алады. Одан әрі қарай ұлпалардың арасында индукциялық байланыс туады, яғни бір ұлпа екінші ұлпаның дамуына әсер етеді. Мысалы, омыртқалыларда хорданың бастамасы гаструляцияның барысында эктодерманың белгілі бір бөлімімен байланысады, соның нәтижесінде эпидермалды клеткалар эктодерманың басқа бөлімдері сияқты тері эпителийіне жіктелмейді, нерв жүйесіне жіктеледі. Геннің әрекеті Геннің қызметі өзгерсе, алдымен зат алмасу содан соң фенотип өзгереді. Мұны микроорганизмдерге жүргізілген тәжірибеден көруге болады. Мысалы, нейроспорада болатын никотин қышқылының синтезделуіне зерттеу жүргізгенде оның кейбір мутантты формаларының бұл қышқылды синтездеуге қабілетті болмайтындығы анықталған. Ал никотин қышқылы синтезінің өзі бірнеше кезеңнен тұрады. Сонда мутацияның әсерінен соның біреуінде генетикалық кедергі болған. Биохимиялық жітелу әрқашан морфологиялық жіктелудің алдында жүреді. Сондықтан гендер әрекетінің бастапқы биохимиялық кезеңін зерттеу өте күрделі болғанымен ген қызметінің механизмін түсіндірудің бірден-бір дұрыс жолы болып саналады. Морфогенезде әрбір жеке ұлпалар мен органдардың жіктелуі біртұтас система болып саналатын организмнің басқа ұлпаларымен байланысты болады. Сонд








