 2015-06-26
2015-06-26 722
722Приведенная выше классификация гемолитических анемий убедительно свидетельствует, что важнейшими этиопатогенетическими факторами развития гемолиза эритроцитов являются нарушения структуры и функции мембран эритроцитов, их метаболизма, интенсивности гликолитических реакций, пентозофосфатного окисления глюкозы, а также качественные и количественные изменения структуры гемоглобина.
В связи с этим целесообразно кратко остановиться на структурных и метаболических особенностях эритроцитов.
Особенности структурной организации и метаболизма эритроцитов. Мембрана эритроцита представляет собой сложный комплекс, включающий определенным образом организованные липиды, белки и углеводы, которые формируют наружный, средний и внутренний слои эритроцитарной мембраны.
На долю белков приходится примерно 49 %, липидов — 44 %, углеводов — 7 %. Углеводы химически связаны либо с белками, либо с липидами и образуют соответственно гликопротеиды и гликолипиды.
Наружный слой образован гликопротеидами с разветвленными комплексами олигосахаридов, которые являются концевыми отделами групповых антигенов крови. Липидным компонентом наружного слоя являются фосфатидилхолин, сфингомиелин и неэтерифицированный холестерин. Липиды наружного слоя мембраны эритроцита играют важную роль в обеспечении постоянства структуры мембраны, избирательной ее проницаемости для различных субстратов и ионов. Вместе с фосфолипидами холестерин регулирует активность мембранносвязанных ферментов путем изменения вязкости мембраны, а также участвует в модификации вторичной структуры ферментов. Молярное отношение холестерин-фосфолипид у человека и многих млекопитающих равно 0,9. Изменение этого соотношения в сторону увеличения наблюдается в пожилом возрасте, а также при некоторых заболеваниях, связанных с нарушением холестеринового обмена.
Снижение текучести мембраны эритроцита и изменение ее свойств отмечаются также и при увеличении содержания сфингомиелина.
Средний бислой мембраны эритроцита представлен гидрофобными «хвостами» полярных липидов. Липидный бислой обладает выраженной текучестью, которая обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами гидрофобной части бислоя. Интегральные белки, к которым относятся ферменты, рецепторы, транспортные белки, обладают активностью только в том случае, если находятся в гидрофобной части бислоя, где они приобретают необходимую для активности пространственную конфигурацию, поэтому любые изменения в составе липидов эритроцитарной мембраны сопровождаются изменением ее текучести и нарушением работы интегральных белков.
Внутренний, обращенный к цитоплазме слой мембраны эритроцита состоит из белков спектрина и актина. Спектрин является специфическим белком эритроцитов, его гибкие вытянутые молекулы, связываясь с микрофиламентами актина и липидами внутренней поверхности мембраны, формируют своеобразный скелет эритроцита. Небольшой процент липидов во внутреннем слое мембраны эритроцита представлен фосфатидилэтаноламином и фосфатидилсерином.
Одним из важных гликопротеинов является гликофорин, содержащийся как на внешней, так и на внутренней поверхностях мембран эритроцитов. Гликофорин в своем составе содержит большое количество сиаловой кислоты и обладает
значительным отрицательным зарядом. В мембране он располагается неравномерно, образует выступающие из мембраны участки, которые являются носителями иммунологических детерминант.
Строение и состояние эритроцитарной мембраны, низкая вязкость нормального гемоглобина обеспечивают значительные пластические свойства эритроцитам, благодаря которым эритроцит легко проходит по капиллярам, имеющим вдвое меньший диаметр, чем сама клетка, и может принимать самые разнообразные формы.
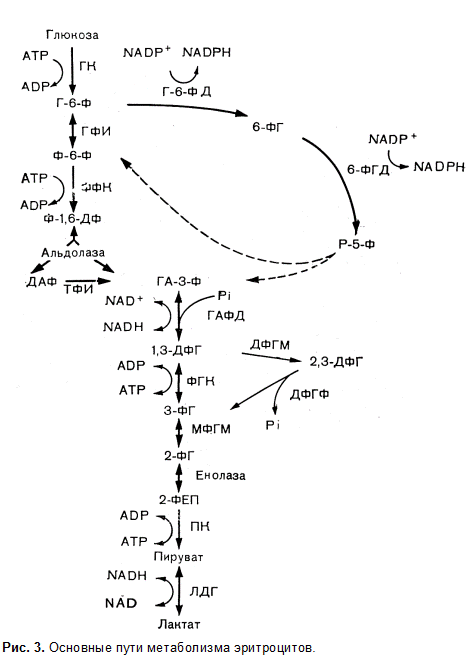
Эритроцит является метаболически активной клеткой и содержит более 40 различных ферментов. Энергетическое обеспечение эритроцита осуществляется за счет утилизации глюкозы в реакциях анаэробного гликолиза. Эффективность гликолиза характеризуется всего двумя молекулами АТФ на одну молекулу глюкозы, однако это небольшое количество энергии обеспечивает эритроциту выполнение всех его функций. Основная доля энергии АТФ расходуется в эритроцитах на транспорт ионов, функционирование АТФазных систем и поддержание электролитного баланса клетки. Макроэргические фосфатные связи АТФ необходимы также и для инициации реакций гликолиза и пентозофосфатного цикла.
Наиболее важные и необратимые реакции гликолиза протекают с участием следующих ферментов: гексокиназы, фосфофруктокиназы и пируваткиназы. Отличительной особенностью гликолиза в эритроцитах по сравнению с другими клетками является выработка значительного количества 2,3‑дифосфоглицериновой кислоты, регулирующей кислородсвязывающую функцию гемоглобина.
Кроме гликолиза в эритроцитах происходит прямое окисление глюкозы в пентозофосфатном цикле, на долю которого проходится 10–11 % всего энергетического метаболизма клетки. Ключевыми ферментами пентозофосфатного цикла являются глюкозо-6-фосфат-дегидрогеназа, 6‑фосфоглюконатдегидрогеназа. В процессе пентозофосфатного окисления глюкозы образуется восстановленная форма кофермента НАДФ, использующаяся для восстановления глутатиона — основного компонента антиоксидантной системы эритроцита.
Главной функциональной группой глутатиона является сульфгидрильная группа, водород которой обеспечивает нейтрализацию органических и неорганических окислителей, действующих на мембрану эритроцита, и защищает липиды мембраны от свободнорадикального окисления.
Вышеизложенное делает очевидным большую значимость для поддержания стабильности эритроцитарной мембраны, интенсивности гликолитических реакций, обеспечивающих образование АТФ и соответственно полноценное функционирование АТФазных систем и трансмембранный перенос ионов, а также состояние пентозного цикла окисления глюкозы и образование достаточного количества НАДФ∙Н2.
В условиях врожденной или приобретенной недостаточности энергообеспечения эритроцитов при нарушении активности гликолитических ферментов, а также ферментов пентозофосфатного окисления глюкозы возникает дестабилизация эритроцитарной мембраны, изменение формы эритроцита и гемолиз.








