 2015-07-04
2015-07-04 9257
9257Для непрерывного переноса ионов натрия и ионов калия через мембрану клетки существует натрий-калиевый обменный насос, который при каждом своем цикле переносит три иона натрия наружу и два иона калия внутрь клетки (рис. 5). Источником энергии для этого процесса является расщепление АТФ, поэтому насос одновременно является ферментом - АТФазой. Специфическими блокаторами натрий-калиевой транспортной системы являются вещества, используемые при лечении сердечной недостаточности - гликозиды, в особенности уабаин. Механизм переноса ионов натрий-калиевым обменным насосом можно представить следующим образом. Внутри каналоподобной структуры насоса расположены места связывания калия и натрия, которые поочередно вступают в контакт с внутриклеточной и внеклеточной средами. Циклическое изменение конформации насоса происходит за счет фосфорилирования и дефосфорилирования его белковой молекулы (присоединение и отщепление остатка фосфорной кислоты). Этот процесс сопровождается изменением сродства мест связывания к соответствующим ионам. Места связывания, направленные внутрь клетки, имеют низкое сродство к ионам калия и высокое - к ионам натрия. Связывание трех ионов натрия ведет к изменению конформации белка, последующему связыванию АТФ и фосфорилированию фермента. Фосфорилирование приводит к дальнейшему изменению конформации, в результате которого места связывания ионов
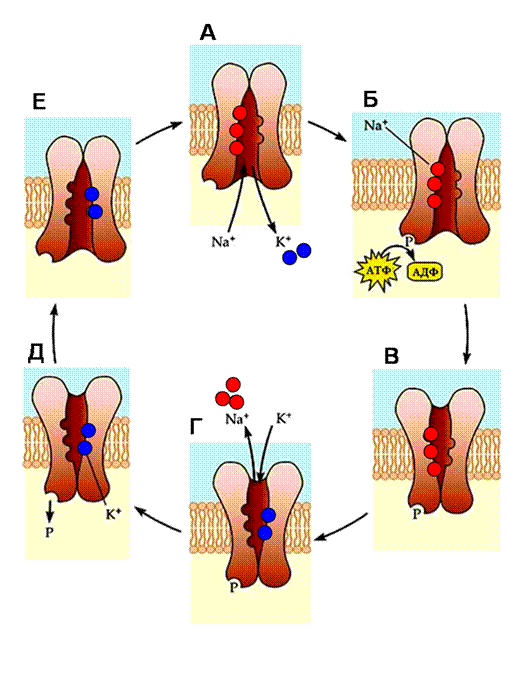
Рис. 5. Перенос ионов натрия и калия через мембрану. Принцип работы натрий- калиевого насоса (натрий- калиевой АТФазы)
(А) Посадочные места, направленные внутрь клетки, обладают высоким сродством к натрию и низким — к калию. Ионы калия, до этого момента связанные с молекулой АТФазы, высвобождаются, в то время как ионы натрия связываются с ней. (Б) Вслед за связыванием натрия происходит связывание молекулы АТФ и фосфорилирование фермента. (В) В результате фосфорилирования в структуре фермента происходят изменения, приводящие к выдвижению посадочных мест во внеклеточную среду. (Г) В таком положении посадочные места обладают низким сродством к натрию и высоким — к калию, поэтому ионы натрия освобождаются во внеклеточную среду, а ионы калия связываются с молекулой АТФазы. (Д) При связывании ионов калия АТФаза дефосфорилируется. (Е) Возврат в первоначальное состояние.
оказываются в контакте с внеклеточной средой. В этом положении места связывания обладают низким сродством к ионам натрия и высоким к ионам калия, поэтому ионы калия замещают ионы натрия. Связывание калия вызывает дефосфорилирование фермента и возвращает насос в первоначальное положение, а ионы калия высвобождаются во внутриклеточное пространство.
Многие функции возбудимых клеток связаны с кратковременным повышением концентрации ионов кальция в цитоплазме, поэтому важной задачей для клетки является поддержание неизменного и очень низкого уровня кальция в покое. В противном случае различные кальций-зависимые механизмы будут активироваться не в ответ на специфическое раздражение, а постоянно. Для поддержания низкой внутриклеточной концентрации ионов кальция наибольшее значение имеют два вида первичного активного транспорта, осуществляемых кальциевыми насосами - АТФазами - непрерывный вывод кальция из клетки через плазматическую поверхностную мембрану и кальциевый транспорт через внутриклеточные мембраны в эндоплазматический ретикулум нейронов (в мышечных волокнах - в саркоплазматический ретикулум) (рис. 6А). Уровень кальция в эндоплазматическом ретикулуме может достигать 400 мкмоль/л, а в саркоплазматическом - 10 ммоль/л. Кальциевый транспортный цикл АТФаз плазматической мембраны в принципе аналогичен циклу работы натрий-калиевого обменного насоса. Он начинается с присоединения иона кальция к месту связывания, расположенному в цитоплазме. Затем происходит фосфорилирование фермента и изменение его конформации, в результате чего ион переносятся во внеклеточную среду. После освобождения кальция молекула АТФазы дефосфорилируется и возвращается в свое первоначальное состояние. Концентрация АТФаз в плазматической мембране нейронов и мышечных клеток довольно низка, поэтому эффективность этой транспортной системы не слишком высока. Кальциевые АТФазы эндоплазматического и
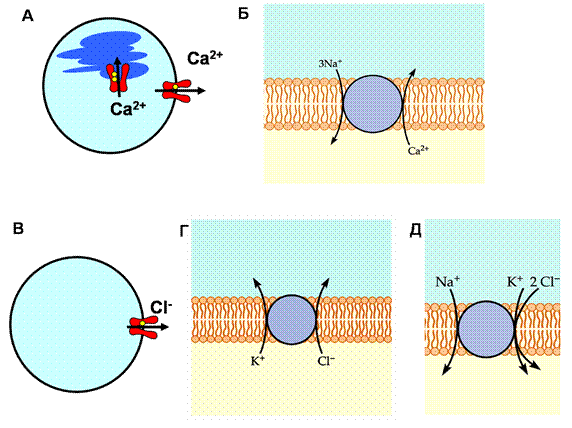
Рис. 6. Транспорт ионов кальция и хлора через мембрану
(А) Внутриклеточная концентрация ионов кальция поддерживается на очень низком уровне, за счет первичного активного транспорта ионов кальция из цитоплазмы в окружающую среду или в эндоплазматический ретикулум (в мышце - саркоплазматический ретикулум). Эффективность второго вида транспорта более значительна, поскольку переносчик за один цикл переносит 2 иона кальция. (Б) Натрий-кальциевый обменник. Выброс одного иона кальция из клетки энергетически сопряжен с входом трех ионов натрия.
Ионы хлора могут как закачиваться в клетку, так и выкачиваться из нее. (В) в некоторых нейронах существует первичный активный транспорт хлора наружу. (Г) Калий-хлорный ко-транспорт осуществляет вывод ионов хлора из клетки за счет энергии выходящего тока калия. (Д) В некоторых нейронах и мышечных волокнах транспорт хлора внутрь клетки производится натрий-калий-хлорным ко-транспортером.
саркоплазматического ретикулумов, за исключением некоторых деталей, не отличаются от кальциевых АТФаз плазматической мембраны. Но поскольку при каждом цикле эта АТФаза переносит два иона кальция, ее мощность значительно больше.
Другой транспортной системой, обеспечивающей низкую концентрацию ионов кальция в цитоплазме, является натрий-кальциевый ионообменник (рис. 6Б). Вместо энергии расщепленной молекулы АТФ он использует энергию перемещения ионов натрия в направлении их концентрационного градиента, то есть внутрь клетки. В результате работы этого ионообменного транспортного механизма внутрь клетки попадает значительное количество натрия. Существуют, по крайней мере, два варианта натрий-кальциевого обмена. Наиболее часто встречаются обменники типа NCX (N a+/ C a2+ e x changer); при их работе происходит перенос одного иона кальция наружу при переносе трех ионов натрия внутрь клетки. Несмотря на то, что сродство NCX к кальцию ниже, чем у кальциевых АТФаз, общая мощность этого транспортного механизма выше приблизительно в 50 раз, поскольку плотность таких молекул в мембране значительно больше. В мембране палочек сетчатки встречается другой тип натрий-кальциевого обменника. В отличие от NCX, он переносит не три, а четыре иона натрия. В работе этого обменника используется дополнительная энергия движения иона калия, также перемещающегося в направлении своего электрохимического градиента (наружу).
Ионы хлора могут как закачиваться, так и выкачиваться из клетки. Хлорных переносчиков очень много - десятки. Хлорные насосы, выкачивающие ионы хлора из цитоплазмы, обнаружены в нейронах мозга (рис. 6В). Тем не менее, большая часть хлора переносится посредством вторичного активного транспорта. Выход ионов хлора из клетки осуществляется хлор-бикарбонатным и калий-хлорным обменниками (рис. 6Г). В некоторых клетках, например, в волокнах скелетной мышцы, аксоне кальмара, происходит активное накопление ионов хлора. Этот транспорт хлора внутрь клетки зависит от внеклеточных концентраций калия и натрия и блокируется широко применяемым в медицине мочегонным лекарственным препаратом - фуросемидом (рис. 6Д).
Для анализа молекулярной структуры мембранных ионных транспортных систем были использованы методы биохимии, молекулярной биологии, генетики и клонирования. В настоящее время мы достаточно много узнали об их строении. Оказалось, что они представляют собой очень крупные белки, аминокислотная последовательность которых в определенных участках спирализуется. Такие спиральные сегменты встраиваются в мембрану, несколько сегментов объединяются в более крупные конгломераты – так называемые мембранные домены. Из них образуются субъединицы белков. При стыковке нескольких субъединиц образуется переносчик.
Большинство переносчиков имеет от 10 до 12 трансмембранных сегментов, которые, как полагают, образует каналоподобные структуры (рис. 7). Перенос веществ через мембрану осуществляется путем попеременного выдвижения посадочных мест во внутриклеточную и внеклеточную среды. Так, натрий-калиевая АТФаза состоит из двух субъединиц: α-субъединица, молекулярная масса которой составляет приблизительно 100 кД, отвечает за ферментативную активность насоса и содержит все места связывания субстрата; меньшая по размеру (35 кД) β-субъединица содержит несколько внеклеточных мест гликозилирования и также необходима для функционирования насоса, однако в чем именно состоит ее роль, неизвестно. Семейство кальциевых насосов плазматической мембраны и эндо- и саркоплазматического ретикулумов представляют собой белковые молекулы с одиночной полипептидной цепочкой молекулярной массы порядка 100 кД, структура которой напоминает α-субъединицу натрий-калиевой АТФазы с увеличенным цитоплазматическим сегментом. В отличие от натрий-калиевой АТФазы, они не нуждаются в β-субъединице для осуществления своей ферментативной активности.
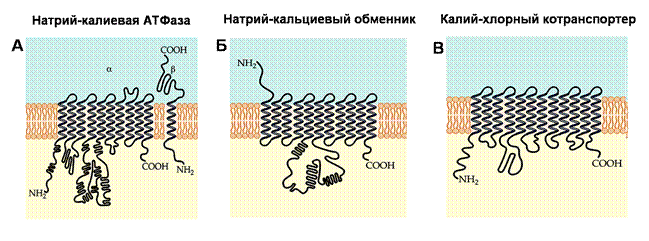
Рис 7. Молекулярная структура транспортных молекул
(А) Натрий-калиевая АТФаза состоит из α-субъединицы, имеющей от 8 до 12 трансмембранных спиральных сегментов, и меньшей по размеру β-субъединицы, пронизывающей мембрану лишь один раз. (Б) Натрий-кальциевые обменники имеют 11 трансмембранных сегментов. (В) Калий-хлорные ко-транспортеры имеют 12 трансмембранных сегментов.

