2015-07-04
2015-07-04 1618
1618У Arabidopsis криптохромная система представлена двумя белковыми комплексами: CRY 1 и CRY 2. У папоротника Adiantum capillus-veneris обнаружено 5 генов, кодирующих различные криптохомы. Судя по последовательности нуклеотидов, белки криптохромов родственны
ДНК-фотолиазам. Эти ферменты позволяют репарировать сшивки, возникающие в ДНК вследствие облучения ультрафиолетом. ДНК-фотолиаза несет две хромофорные группы: флавин и птерин (или деазафлавин). Восстановление ДНК фотолиазами возможно только в присутствии света.
Криптохромы обладают консервативным участком с двумя карманами для хромофорных группировок и дополнительным участком на С-конце. По-видимому, именно С-концевой домен приводит к изменениям свойств фотолиазы и превращению в фоторецептор (рис. 7.31).
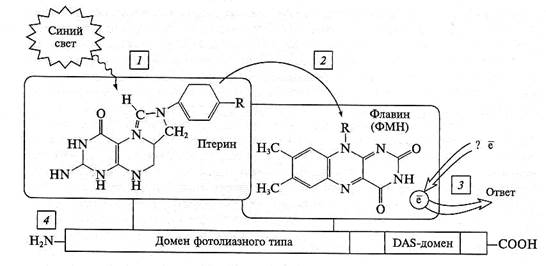
Рис. 7.31. Структура криптохрома:
1 — получение кванта синего света птериновым ядром; 2 — передача возбуждения на ядро флавина; 3 — ответ в форме редокс-реакции; 4 — функциональные домены белковой части криптохрома
Криптохром не может непосредственно восстанавливать ДНК, но первичные акты захвата света у него такие же, как у фотолиазы. Птерин (метенилтетрагидрофолат) функционирует как светособирающий хромофор. При попадании «синего кванта» света возбуждение передается в «реакционный центр» криптохрома — на молекулу ФАД. Наличие двух хромофоров объясняет тот факт, что физиологический спектр действия криптохрома совпадает как со спектром флавинов, так и со спектром птеринов. ФАД в составе криптохрома 1 находится в стабильной семихинонной форме (он частично восстановлен и легко отдает электроны). Семихинон ФАД поглощает не только в синей, но и в зеленой части спектра. Таким образом, физиологический ответ вызывает не только синий, но и интенсивный зеленый свет.
Получив квант света, электрон флавинового ядра возбуждается, т. е. переходит на более высокую орбиталь, и редокс-потенциал резко изменяется. Возбужденный флавин — сильный восстановитель. В настоящий момент существует три гипотезы работы криптохрома. 1. Происходит самовосстановление молекулы криптохрома с изменением ее конформации, что активизирует вторичные мессенджеры. 2. В систему передачи сигнала вовлечен промежуточный редокс-партнер. 3. Партнеру передается не электрон, а возбуждение квантом синего света, и этот квант приводит в действие еще одну хромофорную молекулу. Если эффективного взаимодействия не состоялось, возбужденное флавиновое ядро теряет энергию (флуоресцирует).
На свету криптохром 1 (CRY 1) связывается с фитохромом А, и криптохром фосфорилируется. Очевидно, без фосфорного остатка криптохром не активен. Фитохром А, получив из окружающей среды сигнал о том, что «свет в принципе есть», активизирует систему синего света. Криптохром in vitro взаимодействует с некоторыми фосфатазами, изменяя их активность. Возможно, каскад фосфорилирования/дефосфорилирования — один из механизмов передачи сигнала. Криптохром локализован в ядрах, что неудивительно: он очень похож на ДНК-фотолиазы, которые, по определению, имеют внутриядерную локализацию. Предполагают светозависимый транспорт криптохрома через ядерную мембрану. Вторичные мессенджеры криптохромной системы также располагаются в ядре и участвуют в регуляции экспрессии генов.
Криптохром 2 пока еще недостаточно изучен. Белковая часть молекулы на N-концевом участке похожа на криптохром 1 (соответствует доменам, связывающим хромофоры), отличаются только С-концевые участки. Белок CRY 2 связывает ФАД. При мутациях cry 2 повреждаются ответы на синий свет низкой интенсивности, а если нарушена работа криптохрома 1, то растения не реагируют на высокую интенсивность синего света. Как и в системе фитохромов, у криптохромов есть разделение по функциям: криптохром 1 дает «приблизительный» сигнал (есть много света в синей и соседней зеленой областях), а криптохром 2 сигнализирует более точно (низкая интенсивность синего).
Благодаря спектральным свойствам дополнительного пигмента — птерина, криптохромы поглощают кванты не только синей области, но и из ближнего ультрафиолета (UV-A), поэтому криптохромы рассматривают как возможные УФ-фоторецепторы. Одним из механизмов, защищающих организм от жесткого излучения, является биосинтез антоцианов и каротиноидов. Неудивительно, что их синтез контролируется криптохромами. От криптохромного сигнала зависит экспрессия генов халконсинтазы, халконизомеразы, дигидрофлавонолредуктазы и других ферментов биосинтеза антоцианов. Заметим, что для биосинтеза антоцианов необходимы как фитохромная, так и криптохромная системы.
Криптохромная система участвует в «настройке» внутренних часов в растении и получении фотопериодического стимула, однако она не является главной. Интересно, что у животных (мыши, дрозофилы) также обнаружены криптохромы. У животных криптохромы играют основную роль в фотопериодической регуляции сезонных форм поведения. Криптохромный сигнал замедляет рост гипокотиля (или междоузлий) на свету, помогает фитохрому контролировать процессы деэтиоляции. Обе фоторецепторные системы, действующие в ядерном компартменте, — криптохром и фитохром — в заметной мере дополняют друг друга. Пути передачи сигнала от этих систем, по-видимому, сходятся и взаимодействуют, что приводит к интеграции информации о физиологически важных областях спектра — синей и красной.