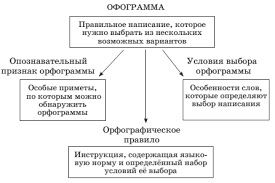
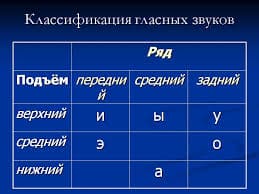
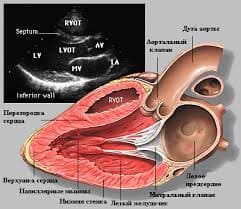
2015-10-16
2015-10-16 952
952За более чем 100-летний период разностороннего изучения популяций и сообществ накоплено огромное количество фактов. Среди них - большое число, отражающих случайные или нерегулярные явления и процессы. Но не меньше и таких, которые после их систематизации обнаруживают проявления устойчивого порядка в существовании популяций и их взаимодействии с внешними процессами и явлениями. Отдельные аспекты таких высокоупорядоченных связей, отношений и процессов, отражающих внешнюю и внутреннюю информацию популяций, формализованы в правила и принципы популяционной экологии - "популяционные законы", отмеченные авторством соответственно приоритетам. Н.Ф.Реймерс насчитывает более 20 таких законов, хотя универсальных и несводимых друг к другу среди них не более десятка.
Некоторые из законов действительно являются отражением порядка, предопределяющего устойчивость популяций в экосистемах. Другие же отражают устойчивость динамических характеристик как бы "задним числом".
Правило объединения в популяции (С.С.Четвериков, 1903): индивиды любого вида живого всегда представлены не изолированными отдельностями, а их определенным образом организованными совокупностями. Формулировка правила, данная Реймерсом, не вполне соответствует содержанию явления, охарактеризованного Четвериковым. Популяция как совокупность, как множество формируется не объединением разрозненных особей, а в процессе воспроизводства пространственно дисперсного видового живого вещества.
Правило колебаний численности: никакая популяция не находится в состоянии уравновешенности числа особей; обязательно помимо сезонных изменений численности возникают периодические флуктуации, обусловленные внешними по отношению к популяции факторами, и осцилляциями, связанные с собственными (внутренними) динамическими изменениями популяции. Правило сформулировано на эмпирическом материале и не требует дополнительных пояснений.
Принцип минимального размера популяций. В связи с тем, что любая популяция обладает строго определенной генетической, фенотипической, половозрастной и другой структурой, она не может состоять из меньшего числа индивидов, чем необходимо для стабильной реализации этой структуры и устойчивости популяции к факторам внешней среды. Минимальный размер представляет собой видоспецифичную константу, а часто - популяционно-специфичную. Снижение численности за этот предел ведет к инбридингу, нарушению механизмов воспроизводства, другим структурным и функциональным аномалиям и в итоге - к вымиранию популяции. Восстановление популяции естественным путем возможно от значений численности выше пороговой, а при численности ниже пороговой это возможно лишь искусственным путем и при условии сохранения типичного сообщества, в которое входила популяция.
Правило популяционного максимума Ю.Одума. Основывается не только на логической посылке, но также на сочетании стремления популяционного живого вещества к максимальной экспансии, встречном давлении смежных популяций и распространения условий, в которых существование популяции является не только возможным, но и необходимым. Численно популяционный максимум равен произведению верхней пороговой величины плотности (за которой начинаются процессы, порождаемые ресурсной недостаточностью) и потенциально пригодной для существования популяции среды как комплекса абиотических и биотических факторов. Полагая, что популяция стремится максимально заполнить доступную среду, хотя никогда не достигает видоспецифичного репродуктивного потенциала, в качестве механизма этого популяционного давления выделим Принцип максимального воспроизводства. Его содержательная сторона состоит в том, что любой популяции присуще внутреннее стремление произвести больше потомков, чем допустимо в реальном сообществе. За счет этого создается как бы слабое давление на смежные популяции, позволяющее до предела заполнить среду, занимаемую живым сообществом.
Численность реальных естественных популяций, не испытывающих сильных или устойчивых отрицательных или положительных антропогенных воздействий, колеблется между порогами максимума и минимума и если достигает этих значений, то лишь на короткое время и при нарушении биоценотических механизмов регуляции.
Принцип минимума и правило максимума получают объяснение в теории Х.Г.Андерварты - Л.К.Бирча (1954), а сама эта теория в расширенном толковании может быть представлена как Правило лимитов популяционной численности: диапазон колебаний численности в нормальных условиях не выходит за нижний порог, нарушающий механизмы воспроизводства, и верхний порог, приводящий к истощению доступных ресурсов и условий для размножения. Правило лимитов имеет и другие формулировки (Н.Ф.Реймерс): "Правило количественной константности популяционного населения" и "Правило максимума размера колебаний плотности популяционного населения". Раскрывая содержательную сторону предлагаемых правил, Реймерс обращает внимание на приведенные в работе Ю.Одума примеры характерных для различных животных порядковых значений плотности популяции. Учитывая их, можно несколько расширить толкование "Правила лимитов" добавлением:... при этом существуют видоспецифичные значения уровней пороговых величин плотности популяции.", что делает правило универсальным.
Принцип К.Фридерихса (1927) экосистемной регуляции численности популяции: регуляция численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида. Реймерс поясняет, что экосистема формирует потенцию роста численности популяции, заложенную генетически, но сама экосистема может в определенных обстоятельствах давать сбои и тогда численность популяции дает вспышки (взрывной рост). С таким комментарием нельзя согласиться полностью. Если мы понимаем экосистему как живую систему некоторого уровня, включающую биотические и абиотические элементы в их разнокачественных состояниях, то определение "нормативного" и "сверхнормативного" минимума и максимума связано с нашими представлениями об устройстве и функционировании экосистемы на основе регулярно повторяющихся явлений и процессов. Но, в случае взрывного роста, популяция, тем не менее, остается частью этой же самой экосистемы, и частью неотделимой. Здесь невозможно абсолютно разделить внутрипопуляционные, межпопуляционные - внутриэкосистемные биотические и внутриэкосистемные абиотические факторы. Поэтому там, где мы находим якобы нарушения экосистемных механизмов регуляции, лежит лишь очередной пороговый переход на более жесткие экосистемные механизмы регуляции, так или иначе приводящие уровень численности популяции к среднеисчисляемым значениям.
Правило пищевой корреляции (В.Уини-Эдвардс): в ходе эволюции сохраняются только те популяции, скорость размножения которых скоррелирована с количеством пищевых ресурсов среды обитания. Это правило представляет частный случай предыдущего и сводимо к нему, поскольку зависимость от пищи - есть лишь один из аспектов зависимости популяции от экосистемы и отражает один из механизмов регуляции численности популяции внешними факторами. Комментируя это правило, Реймерс подчеркивает, что скорость размножения всегда ниже максимально возможной и поэтому популяция всегда имеет запас пищевых ресурсов. Он относит такое явление к эволюционно выработанной преадаптации более низкой плотности популяции, чем допускают наличные кормовые возможности. К сказанному можно добавить, что в пищевых цепях каждое последующее звено в норме "снимает" только избыточную биомассу, продуцируемую предыдущим звеном, оставляя основную производящую часть биомассы. Таким образом, в процессе эволюции должна вырабатываться не односторонняя преадаптация популяции к общему наличному запасу корма, а двусторонняя взаимоадаптация смежных в пищевой цепи популяций, выражающаяся в соответствии избыточно продуцируемой и потребляемой биомассы. Данный вывод имеет непосредственный выход на Правило пирамиды энергий, или Правило 10% Р.Линдемана: с одного трофического уровня экологической пирамиды переходит на другой, более высокий ее уровень, в среднем, около 10% поступившей на предыдущий уровень экологической пирамиды энергии.
Принцип Д.Лэка (1954): среднее количество яиц, откладываемых птицами, эволюционно ограничено возможностями выращивания выведшихся из них птенцов. Этот принцип полностью идентичен по содержанию Правилу пищевой корреляции, но уже его по форме.
Теория Дж.Кристиана - Д.Дейвиса (1950-68) объясняет регуляцию численности популяции социальными механизмами, то есть внутренними механизмами, в которых задействованы эндокринные реакции особей. Частным проявлением этой теории является Правило стрессогенного увеличения надпочечников у высших позвоночных животных: в условиях перенаселенности у них увеличиваются надпочечники, что ведет к повышению агрессивности, снижению репродуктивного потенциала, устойчивости к заболеваниям и другим последствиям. Одновременно данное правило объясняется Теорией общего адаптационного синдрома Г.Селье.
Правило сохранения видовой среды обитания (В.Уини-Эд-вардс, Н.Ф.Реймерс, 1950-е - 1960-е) является суммарным производным от правил популяционного максимума и пищевой корреляции. Его формулировка: - животные в норме не могут абсолютно разрушительно действовать на среду своего обитания, поскольку такое разрушение лишает их собственной основы жизни. В то же время, популяция может осуществлять компенсаторную количественную реакцию на изменения во внешней среде, ослабляя или усиливая воздействие на нее, и тем самым участвуя в экосистемных механизмах ее стабилизации. Содержательно это положение соответствует Принципу А.Николсона.
Для каждого вида характерен свой особенный тип онтогенеза, включающего последовательность различных возрастных и фазовых состояний, жизненных форм. Отдельные видовые популяции, входя в конкретные биогеоценозы, отличающиеся хотя бы по некоторым параметрам, приобретают более или менее явные компенсаторные изменения, которые могут быть выражены и в особенностях строения, и в наличии и длительности отдельных возрастных состояний. Развитие одноименных особей в популяции может проходить синхронно и асинхронно. Среди них, обычно, существует более или менее устойчивое разделение на половые типы с их особенностями онтогенеза и способа жизнедеятельности. Все это создает в популяции некоторую, закономерно изменяющуюся во времени мозаику из определенных половозрастных групп. Отсюда вытекает "Принцип сохранения специфического типа динамики половозрастной структуры популяции".
На известных принципах "территориальности" и "скопления особей", определяющих, с одной стороны, наличие индивидуальных и групповых территорий, а с другой стороны, агрегацию в стаи, стада, колонии, повышающих выживание групп особей по сравнению с одиночными особями, устанавливается более общее Правило топографического, или популяционного кружева ареала. Оно заключается в том, что популяция заселяет пространство неравномерно, оставляя "пустые" места, которые непригодны для ее жизни, и, распадаясь на экологически разнородные микропопуляции, каждая из которых приурочена к определенному местообитанию.
Принцип стабилизации экологических ниш, или Принцип биоценотической коэволюции: эволюция популяций внутри сообщества и эволюция этого сообщества скоррелированы таким образом, что каждый вид устойчиво сохраняет в этой природной системе свое функциональное место (то есть экологическую нишу) до тех пор, пока внешние силы (воздействия надсистем или аналогичных систем) не изменят существующего баланса. Даже небольшое устойчивое в длительности изменение соотношения между (биотическими, абиотическими) элементами вызывает образование "пустот" между частично перекрывающимися эконишами - основы для внедрения или образования новых элементов. В других случаях может происходить разделение емкой экониши на две или более субниши, позволяющее более эффективно и полно использовать имеющиеся экосистемные ресурсы. Это положение известно как Принцип В.Людвига и его можно рассматривать в качестве частного случая "принципа стабилизации".