 2015-10-13
2015-10-13 1997
1997Влияние смены циклов природной среды на популяции осуществляется через изменение емкости угодий, пригодных для жизни (степень обилия, качества, разнообразия корма и убежищ), изменение биоценотических связей, интенсивности размножения и т.д.
Изменение условий существования популяций, влияя на их репродуктивность и численность, включает в действие факторы, зависимые от плотности популяции (влияние хищников, паразитов, развитие стресса, болезней, эпизоотии), ведет к перестройке их эндокринного комплекса и генетической структуры и др. Следовательно, смена очередной фазы природного цикла, приводя к смене условий существования популяций и влияя на их численность, формирует систему авторегуляторных механизмов динамики, специфичных для разных фаз популяционного цикла.
В циклической изменчивости природной среды каждый вид занимает свою временную экологическую нишу, которая благоприятна для роста численности популяции и развития вспышки массового размножения. Ее продолжительность определяется длительностью соответствующей фазы природной среды. Некоторые популяции одних видов опосредованно через циклы природной среды реагируют ростом численности на высокие показатели солнечной активности, а другие - на низкие. То есть, виды, реагирующие опосредованно через среду обитания на высокие показатели солнечной активности, находят оптимум условий существования при фазе природного цикла, соответствующего этому уровню активности Солнца, и наоборот (Максимов, 1984). В динамике численности населения зверей выделяют три типа (Н. Наумов, 1948; Северцов, 1941):
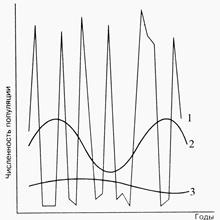 1. Скачкообразное движение, характерное для мелких грызунов и землероек с очень быстрым оборотом стада (рис.3);
1. Скачкообразное движение, характерное для мелких грызунов и землероек с очень быстрым оборотом стада (рис.3);
2. Неустойчивое движение с резкими изменениями численности, характерное для белки, лисицы, песца и др.;
Рис. 3. Типы динамики численности животных: 1 – скачкообразный, 2 – неустойчивый, 3 – устойчивый (Северцов, 1941)
3. Устойчивое движение населения с медленными, растянутыми во времени и плавными подъемами и сокращениями численности (слоны, носороги, киты, лоси, рукокрылые и др.).
Различают фазу подъема численности населения (размножение резко превышает смертность), пик численности (высокая плотность населения тормозит размножение и выживание молодых), фазу падения численности (сложившиеся условия вызывают большую смертность от голода, неблагоприятной погоды или заболеваний) и фазу депрессии (снижение численности прекращается, начинают восстанавливаться условия для ее нового подъема) (Н. Наумов, 1958). У всех видов на фазе подъема численности общей особенностью является закономерное изменение условий существования и, как следствие, сменяются ведущие лимитирующие факторы движения населения.
У ряда видов животных существуют 10, 12, 30, 60-летние и т.д. циклы изменения численности. Многие мелкие млекопитающие, кроме 10-12-летней цикличности или в отсутствии ее, имеют кратные этой периодике гармонические составляющие численности с периодом 6 лет и 3-4 года. Кроме того, указанная цикличность может сама по себе служить синхронизатором ритмов с большими периодами (5-6-летняя периодичность «наводит» 10-13-летний цикл). Например, серые полевки имеют на спектрах своей численности пики, соответствующие 3-летней цикличности.
Массовые виды грызунов (лемминги, зайцы, белки и др.), взаимодействуя со своей кормовой растительностью, проявляют изменения численности, аналогичные взаимодействию хищник-жертва. При падении численности грызунов (жертв) сокращается численность и хищников. Регулярность циклов объясняется тем, что действие основного фактора – взаимодействия хищник-жертва – лишь в незначительной степени осложняется влиянием других факторов, а тенденция к независимым изменениям численности в разных районах устраняется миграциями.
Скачкообразное движение численности (рис. 3, п.1).Массовые виды мелких грызунов (мыши, полевки, лемминги и др.) отличаются наиболее высокой плодовитостью среди всех млекопитающих, но самой низкой социальной организацией и очень большими ее колебаниями в разные сезоны и годы. Размножение сокращается, размер выводка уменьшается, плотность населения и площадь обитания резко сокращаются при неблагоприятных условиях.
Животные максимально быстро для воспроизводства поголовья используют возникающую благоприятную обстановку - обильный урожай корма, теплая затяжная осень, мягкая зима и т. п. При интенсивном подъеме численности молодняк в короткий срок занимает потерянные при вымирании участки, и распределение популяций грызунов из пятнистого, спорадичного, становится сплошным или почти сплошным. То есть в фазу пика численности территория и ее ресурсы используются ими наиболее полно, резко увеличивается количество заселенных биотопов. Кроме того, часть животных мигрирует в разных направлениях, благодаря чему ареал временно расширяется, как это бывает у норвежского лемминга, появляющегося на Скандинавском и Кольском полуострове на десятки километров южнее обычной границы распространения (здесь возникают временные колонии иммигрантов).
При интенсивном размножении численность полевок и мышей за короткий период увеличивается в сотни раз, так как молодые зверьки становятся половозрелыми, еще не достигнув полного роста, и в свою очередь приносят один выводок за другим. В период массового размножения полевок зверьки десятками в течение всего дня то и дело попадаются на глаза. Множество дорожек, кучек нарытой земли, поврежденных растений говорит об их интенсивной деятельности. Размножившиеся степные пеструшки в Средней Сибири и полевки Брандта в Забайкалье и Монголии полностью уничтожали растительный покров пастбищ и сенокосов, крайне затрудняя выпас домашних стад.
В период массовых размножений, когда численность полевок и мышей достигает наивысшего уровня, репродуктивный процесс начинает затормаживаться и плодовитость резко снижаться. Снижается плодовитость и у лисиц при ее сильной перенаселенности.
Другая причина, сдерживающая размножение грызунов в годы высокой численности, - глистные инвазии, распространение которых (экстенсивность и интенсивность инвазированности зверьков) увеличивается при высокой плотности их населения. Известно, что продукты жизнедеятельности гельминтов способны оказывать токсическое действие на организм хозяина и угнетать его половую продуктивность вплоть до появления полной стерильности.
Таким образом, падение численности населения мелких грызунов после вспышки размножения обычно вызывается сочетанием целого ряда факторов как эндогенного, так и экзогенного происхождения.
При свойственной мышам и полевкам высокой плодовитости их население может достигнуть высокого уровня численности за короткий ряд лет. При малой стойкости против условий, вызывающих смертность, и, следовательно, малой продолжительности жизни столь же быстро происходит падение численности. Помимо этих частых, местных подъемов численности животных на небольших площадях в движении населения степных и полевых грызунов различают массовые размножения, происходящие не чаще одного раза в десятилетие и охватывающие огромные территории («большие волны»). В эти грандиозные массовые размножения вовлекаются многие виды грызунов. После очередного сокращения численности полевки и мыши сохраняются обычно в немногих изолированных участках-«резервациях», или «стациях переживания».
Неустойчивое движение численности (рис.3, п.2).Воспроизводительная система узкоспециализированных животных-семеноедов очень чувствительна к измененям кормовых условий. Даже среди видов, питающихся грубыми растительными кормами, способных усваивать не только свежую зелень, но и засохшую, выгоревшую траву, сухие побеги и т. п., установлены случаи значительных изменений численности из-за с недостатка или обилия пищи.
В движении численности обыкновенной белки урожай плодов (семян) играет роль важнейшего внешнего толчка к массовому размножению. Малая плодовитость белки (по сравнению с мышами и полевками) обусловлена ее большей стойкостью и относительно высокой продолжительностью жизни: в природных условиях часть белок доживает даже до 6-7 лет. Для мелких мышей в урожайный на семена год характерно длительное осенне-зимнее размножение, начинающееся обычно после опадения на землю желудей дуба, орешков бука, липы и др. и продолжающееся до начала лета следующего года включительно. Белки же при высоком урожае семян ели успевают за счет высококалорийного свежего корма принести лишь один выводок в конце лета того же года (конец августа-сентябрь). В основном же используют его в теплую половину следующего года, поэтому и подъемы их численности всегда отстают на один год по сравнению с урожайными годами лесных пород.
Вызываемое недостатком семян изменение рациона белки (использование вынужденных и балластных кормов) обусловливает истощение организма животного, заболевания, откочевки и в конечном итоге - массовую гибель. Особенно много истощенных белок погибает в морозные месяцы зимы и при поздне-осенних миграциях, что сильно сокращает количество производителей перед началом гона. Длительное использование биологически неполноценного корма сильно сказывается на половой функции белки, вызывая резкое снижение плодовитости и даже полное прекращение размножения. В итоге после неурожая семян шишек хвойных деревьев, а особенно после двух последовательных неурожайных лет, численность белки резко сокращается. В зависимости от чередования семенных и неурожайных лет основной кормовой породы «урожаи» поголовья белки повторяются с промежутками в 3, 4, 5, 6 лет. Даже в пределах одной области или республики (например, Якутии) разные ее районы могут существенно отличаться в частоте «урожаев» белки, в их величине и т. д. Это обусловливается особенностями местных физико-географических условий, наличием лесных площадей, средними размерами урожая семян, продолжительностью периода размножения белки и т. п.
Наблюдения в тундрах России показали, что «урожаи песцов» на северо-востоке Сибири повторяются через каждые 3-4 года. Для севера Канады «урожайные» годы повторялись через 3-5 лет, в среднем - 3,9 года. Одновременным урожаем или неурожаем охватывается только треть или четвертая часть огромного протяжения тундр. Повсеместно высокая численность песцов обусловливается массовым размножением леммингов.
Осенью в год высокой численности песцов, часто совпадающий с вымиранием леммингов, начинаются массовые перекочевки хищников. Среди мигрантов распространяются эпизоотии, наносящие огромный урон поголовью. Преобладают случаи поражения вирусом «дикования» (название болезни). Ездовые собаки, лисицы и волки тоже вовлекаются в эти эпизоотии. «Дикование» некоторыми симптомами напоминает бешенство. Большинство песцов умирает в первую же зиму после начала эпизоотии. Количество трупов, вытаивающих весной с сохранившимся хорошим мехом, нередко бывает огромным.
Движение численности лисицы в основном сходно с описанным для песца, но имеет и существенные отличия, обусловленные необычайно широким ее распространением и высокой экологической пластичностью. Популяции лисиц, занимающие разные природно-географические зоны, характеризуются каждая своими особыми вариантами движения численности, черты которых обусловлены как естественными биотическими и абиотическими факторами, так и влиянием хозяйственной деятельности человека.
В средней и степной полосе страны, где в населении мелких грызунов господствуют обыкновенные и общественные полевки и степные пеструшки, подъемы и падения их численности повторяются обычно с промежутками в 2-3 года, вызывая соответствующие, но более сглаженные, изменения числа лисиц. Картина осложняется влиянием «больших волн» массового размножения грызунов, повторяющихся с интервалами в 9-10 лет. Наличие обусловленных «большими волнами» и почти синхронных с ними размножений лисиц служит доказательством реальности существования вспышек размножения грызунов с 10-летней цикличностью. Помимо снижения плодовитости длительный недостаток продуктивного корма вызывает увеличение смертности взрослых лисиц: часть их погибает от голода, часть от болезней, усиливающихся в связи с истощенностью животных.
В зоне тайги глубокий и рыхлый снежный покров ограничивает доступность кормов у лисицы, и ставит ее в тесную зависимость от зайца-беляка, подверженного резким колебаниям численности. Это в основном и обусловливает движение населения таежных лисиц по годам. В тайге Сибири и Северной Америки высокая численность лисиц отмечается с промежутками в 8-10 лет.
Хорошее размножение соболя в Зауралье обусловливается обилием рыжих полевок, серых полевок и лесных леммингов. В годы их обилия (продуктивного корма соболей) 90-100% соболюшек участвуют в гоне и оказываются оплодотворенными. Число эмбрионов колеблется от 2 до 6, а в среднем равняется 4-4,5. В год ухудшения запасов продуктивного корма число оплодотворенных самок было 70%, а среднее число эмбрионов было 3,2. Колебания плодовитости несомненно оказывают влияние на изменение численности соболей, но, при свойственной виду значительной стойкости, падения и подъемы кривой, показывающие движение их населения, гораздо более плавны и не столь резки, как у грызунов или узкоспециализированных хищников-мышеедов.
Урожайные и неурожайные на соболя годы повторяются с интервалами в 11 лет (Фолитарек, 1947). В Северной Америке у близкой к соболю американской куницы подъемы численности повторяются в среднем через 9,5-10 лет.
К группе видов с неустойчивой численностью населения принадлежат и зайцы, в частности наши русак и беляк. У беляка в разных частях огромного ареала движение населения имеет некоторые местные отличия. По своей неприхотливости в отношении питания беляк напоминает лесных оленей, питающихся большую часть года веточным кормом, и некоторых серых полевок. Состояние и доступность корма в движении численности популяции беляка играют второстепенную роль. Беляки оседлы, массовые перекочевки бывают у них исключительно редко.
В таежной полосе численность беляка колеблется в 4-6-кратных пределах, в лесостепной в 3-4-кратных. У северо-восточной границы ареала, в отдельных районах Якутии количество беляков за 5-10 лет изменяется в сотни и даже в 2500 раз (С. Наумов, 1961). Здесь продолжительность циклов (от одного пика до другого) составляет около 12 лет. В Западной Сибири она равна 8-10 годам, на Европейском Севере - 9-11, в Ленинградской области – 5-7, в Московской – 4-5 и т. д. Следовательно, в областях с более суровыми природными условиями нужно большее число лет, чтобы сохранившиеся производители могли обеспечить достаточный рост популяции.
Выяснено, что плодовитость беляков меняется по годам, причем это обусловливается изменением величины выводка, тогда как процент самок, участвующих в размножении, остается постоянным (несомненное влияние относительно равномерной обеспеченности кормом в разные годы). Основная причина колебаний плодовитости - интоксикация организма самок, возникающая на почве глистных и протозойных инвазий. Она уменьшает вероятность имплантации оплодотворенных яиц, вызывает резорбцию эмбрионов. Но изменение плодовитости по годам не единственная причина колебаний численности беляка; она только усиливает несоответствие между рождаемостью и смертностью. Последняя в фазу падения численности велика и в основном обусловлена гибелью зайцев от болезней и деятельности хищников, уничтожающих много ослабевших зайцев. То есть на движение численности беляка условия погоды воздействуют не прямо, а через посредство паразитических организмов. Они вызывают у беляков снижение плодовитости и тяжелые заболевания, нередко кончающиеся смертью наиболее инвазированных особей.
Устойчивое движение численности (рис. 3, п.3).Большинство крупных млекопитающих (копытные, хищные, китообразные, ластоногие и др., а из мелких – рукокрылые)отличается достаточной стойкостью против неблагоприятных внешних влияний и значительной продолжительностью жизни особей. У большинства видов высокая социальная организация жизни и наиболее сложный набор поведенческих актов. Они много времени уделяют воспитанию своих детенышей, поэтому численность их населения изменяется медленно, без резких подъемов и депрессий, так как плодовитость невелика, мало изменяется по годам. Процент смертности тоже держится на относительно постоянном низком уровне. Летучие мыши отличаются очень низкой плодовитостью - самка приносит в год только одного, а у некоторых видов двух детенышей, но продолжительность жизни этих зверьков до 15-20 лет. Численность летучих мышей в пещерах, дуплах, старых зданиях изменяется медленно. Но иногда внезапно животные исчезают, сменяя свои убежища.
Лось в естественных условиях доживает до 13-15 лет и более. В норме плодовитость 1-3, в среднем 1,4 теленка. Смертность наиболее высока среди телят в возрасте до 6-7 месяцев. Взрослые лоси от болезней, стихийных бедствий и хищников страдают относительно мало и погибают в небольшом количестве.
Продолжительность жизни северного морского котика равна 14-15 годам; взрослая самка приносит за год только одного детеныша (двойни очень редки). Гибель котиков велика только в первые месяцы жизни, так как много молодых давят тяжелые секачи при драках на лежбищах, часть умирает от сильной инвазированности нематодамиили от голода в случае гибели матери и т. п. Всего в первые 5 месяцев жизни погибает до 20% молодых «черных» котиков. Позднее в море некоторое число неокрепших животных уничтожают косатки.
У видов с таким ходом размножения и смертности заметные подъемы и депрессии численности возможны только при воздействии каких-то длительно действующих факторов, например, постоянного интенсивного промысла, больших многолетних сдвигов гидрологических условий.

