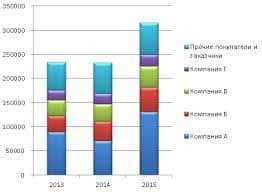
2015-10-13
2015-10-13 2575
2575Закономерности голосообразования и звукового общения у птиц — одно из важнейших направлений в современной орнитологической биоакустике. Исследование вопросов функциональной физиологии голосового аппарата птиц связано с большими трудностями, главным образом в связи с многообразием морфологических типов нижней гортани у различных систематических групп класса (Тереза, 1930; Ames, 1971). В последнее время наиболее перспективным методом изучения голосообразования служит анализ с помощью специальной радиоэлектронной аппаратуры акустической структуры излучаемых птицами звуков. Использование этого метода применительно к раннему онтогенезу дает возможность выявить и возрастные закономерности голоса у птиц.
ПРЕНАТАЛЬНЫЕ СТАДИИ РАЗВИТИЯ ГОЛОСА У ПТИЦ
Становление акустической сигнализации у птиц в эмбриогенезе освещено в литературе чрезвычайно слабо. Основное внимание исследователи уделяли «щелкающим» звукам эмбрионов, как наиболее легко регистрируемым непосредственно перед вылуплением.
Звуковое общение, будучи надежным механизмом связи, широко используется выводковыми птицами, у которых развитие пуховой системы в эмбриогенезе идет опережающими темпами по сравнению с развитием зрения. Микрофонный потенциал улитки куриного эмбриона в ответ на низкочастотные звуки регистрируется на 11-й день инкубации, а электрическая активность сетчатки глаза — только на 18-й день.
Установлению взаимного общения способствует гетерохронное развитие слухового анализатора эмбрионов. Оно обеспечивает максимальную слуховую чувствительность перед вылуплением в диапазонах частот, соответствующих основным энергетическим максимумам в звуковых сигналах родителей и собственной вокализации. Акустическая афферентация на определенных стадиях раннего онтогенеза оказывает непосредственное влияние на развитие слуха, ускоряет процесс освоения высокочастотного диапазона, характерного для собственной вокализации эмбриона. Диапазон воспринимаемых частот птенцов как у выводковых, так и у полувыводковых птиц совпадает со спектральными характеристиками эффективных для соответствующих форм поведения видоспецифических сигналов взрослых птиц, что имеет важное адаптивное значение. Оно состоит в том, что видоспецифическая звуковая сигнализация между эмбрионами и взрослыми птицами обеспечивает синхронизацию вылупления выводка и поддержание стабильности его последующего существования.
Развитие акустической сигнализации у птиц в пренатальном онтогенезе опосредовано становлением легочного дыхания. Первые звуковые сигналы эмбрионов формируются еще до выхода их в воздушную камеру яйца. По времени появления они соответствуют «спонтанному» дыханию, которое осуществляется за счет воздуха полости амниона. В этот же период устанавливается и взаимная акустическая связь эмбрионов и насиживающей птицы. Такое явление отмечено у куликов, на примере большого веретенника, у куриных и пластинчатоклювых.
Начало функционирования звукоиздающих систем у представителей различных систематических групп значительно варьирует. Первые звуковые сигналы эмбрионов представляют собой одиночные писки, разделенные длительными временными интервалами — до 30—60 мин. После выхода эмбриона в воздушную камеру яйца его звуковая активность резко увеличивается, что свидетельствует о появлении истинного легочного дыхания. Интенсивность писков возрастает, их можно слышать даже без вскрытия скорлуповой оболочки яйца, но они по-прежнему разделены длительными паузами — 20— 40 мин. Проклев — появление первых трещин на скорлупе — сопровождается группированием отдельных писков в серии из 2—-3 импульсов. Двигательная активность эмбрионов на этой стадии развития сопровождается интенсивными писками; частота их излучения значительно увеличивается при резких движениях и вибрации яиц.
Длительность паранатального периода (от проклева скорлуповой оболочки до вылупления) коррелируют у птиц с общей продолжительностью инкубационного периода. Обращает на себя внимание короткий паранатальный период у нанду и чомги. Этот парадокс связан с экологией гнездования вида. Сокращение длительности паранатального периода у нанду до минимума является своеобразной адаптацией эмбриогенеза к аридным условиям. Проклев скорлуповой оболочки эмбрионом перед вылуплением приводит к интенсивному испарению влаги, что при длительном паранатальном периоде развития в условиях саванн и полупустынь может достигнуть критической величины и повлечь за собой гибель кладки. В гнездах поганок, напротив, отмечена высокая влажность, обусловленная известными особенностями их «плавучей» конструкции. Длительное пребывание эмбрионов на стадии проклева скорлупы в условиях повышенной (избыточной) влажности также может оказаться для них губительным. В связи с этим, несмотря на раннее включение звукоиздающей системы эмбрионов, длительность паранатального развития чомги сокращена до минимума.
«Щелкающие» звуки занимают особое положение при развитии голоса у птиц. Они сопутствуют легочному дыханию и характерны для эмбрионов. Существует мнение, что «щелкающие» звуки возникают в результате подвижности хрящей трахеи, бронхов или гортани. Как показали исследования, «щелчки» являются вторым типом звуковых сигналов в хронологическом порядке при развитии голоса у птиц в эмбриогенезе. Первые «щелчки» — нерегулярные и низкоинтенсивные — регистрируются у эмбрионов за несколько часов до проклева скорлуповой оболочки. Их ритм не превышает 10 в минуту. Серии, включающие от 10 до 50 импульсов, чередуются с паузами длительностью до 5—15 мин.
Проклев скорлупы и последующая стабилизация легочного дыхания приводят к становлению регулярной и более интенсивно «щелкающей» активности у эмбрионов. Поскольку «щелкающие звуки сопутствуют дыхательным актам, их ритм нарастает вплоть до вылупления, являясь индикатором развития и стабилизации дыхания. По спектрально-временным параметрам они представляют собой короткие (10—30 мс), ритмические широкополосные импульсы. Видоспецифичных характеристик «щелкающих» звуков не обнаружено. Ритм «щелчков» помимо возрастных особенностей эмбрионов находится в прямой зависимости от внешней температуры, что вызвано интенсификацией дыхательных движений. У выводковых и полувыводковых птиц «щелкающие» звуки служат основой для акустической стимуляции эмбрионов, приводящей к акселерации эмбрионального развития и синхронизации вылупления птенцов в кладке.
Переход эмбрионов к дыханию атмосферным воздухом сопровождается ритмической организацией излучаемых звуковых сигналов. Определенные категории из них (сигналы «дискомфорта», «комфорта») имеют функциональное значение в процессе звукового общения эмбрионов и насиживающей птицы. У ряда групп проклев скорлуповой оболочки и стабилизация легочного дыхания эмбрионов резко изменяют спектральную структуру излучаемых сигналов. В целом же переход к излучению «шумовых», или широкополосных сигналов, практически не имеющих выраженной частотной модуляции, имеет место у птиц с «примитивным» типом строения нижней гортани. Примитивный тип строения нижней гортани характеризуется одной парой мышц, а у некоторых видов голенастых (аисты) и бескилевых птиц (эму, нанду, африканский страус) и она подвергается значительной редукции. Развитая нижняя гортань (например, у певчих воробьиных) обусловливает сложность голосовой мускулатуры (8—12 пар); ей свойственна сильная модификация окостеневающих трахеальных колец.
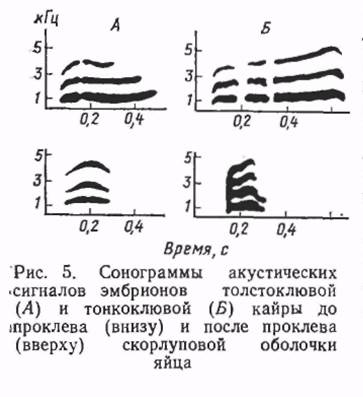
Различна и структурно-динамическая организация сигналов. Эмбрионы толстоклювых кайр способны излучать как отдельные импульсы, так и сигналы трелевого звучания. Трелевая структура сигналов не характерна для пренатального онтогенеза тонкоклювых кайр. Столь раннее и сильное различие акустических сигнальных систем у близких видов кайр обусловлено, по-видимому, ихсовместным гнездованием в колониях. Высокого уровня в гнездовых колониях кайр достигает не только межвидовое, но и индиви- дуальное опознавание в семьях.
Зрелость и сложность акустической сигнальной системы у птиц к моменту вылупления определяются типом развития и видовыми экологическими особенностями. В пренатальном онтогенезе выводковых и полувыводковых птиц формируются все основные категории сигналов: звуки «дискомфорта», «комфорта», «выпрашивания корма» и т. д. Не регистрируются у эмбрионов лишь сигналы тревоги.
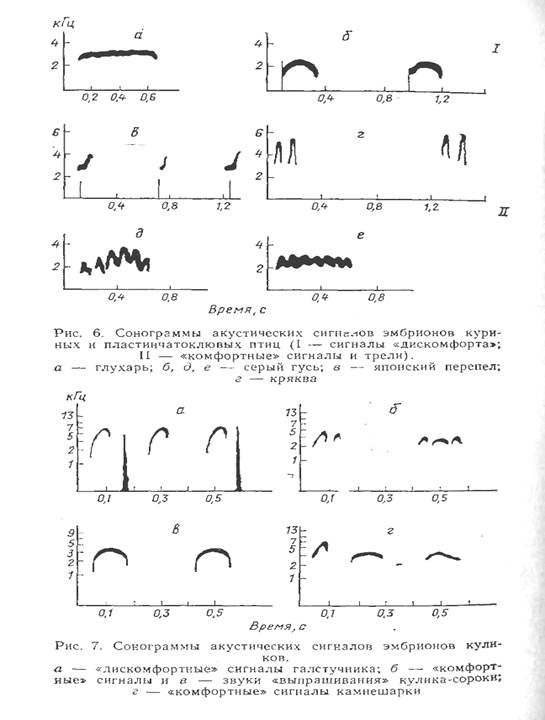
Эмбрионы глупышей (Fulmarus glaclalis) и поморников (сем Stercorariidae) на стадиях предвылупления способны издавать все звуковые сигналы, характерные для взрослых птиц. Сравнительный анализ ювенильной и дефинитивной акустических сигнальных систем у этих видов свидетельствует, что возрастные изменения выражаются главным образом в расширении спектральных границ и увеличении длительности сигналов. Структурная организация звуковых сигналов у эмбрионов и взрослых птиц практически идентична. Таким образом, у трубконосых и поморников тип развития акустической сигнальной системы жестко детерминирован. Все категории звуковых сигналов формируются в пренатальном онтогенезе и по структурной организации являются как бы копиями дефинитивных сигналов. Дальнейшей функциональной дифференциации и структурного усложнения сигналов не происходит.
Перед вылуплением эмбрионы активно отвечают сигналами «дискомфорта» на определенные внешние воздействия: охлаждение, резкие перевороты яиц, встряхивание и т. п. Число импульсов в серии и ритм их излучения не строго фиксированы и определяются, по-видимому, физиологическим состоянием эмбрионов и внешними факторами. Сигналы «комфорта» легко отличимы на слух от «дискомфортных» сигналов и воспринимаются как тихое щебетание или посвистывание. Интенсивность их излучения эмбрионами значительно ниже, чем сигналов «дискомфорта». Сигналы «комфорта» обычно регистрируются в конце «вспышек» двигательной активности у эмбрионов, при согревании охлажденных яиц, их вибрации.
К одной из разновидностей звуков «комфорта» относятся «комфортные» трели. Трели издаются эмбрионами на стадиях непосредственно предшествующих вылуплению. Трели следуют, как правило, в конце серии «комфортных» звуков и завершают ее. Для эмбрионов пластинчатоклювых, куриных, пастушковых и некоторых других видов птиц характерны «сонные» трели как один из вариантов трелевых звуков. Они отличаются от обычных «комфортных» трелей узкополосностью в спектральном отношении, и меньшей длительностью импульсов. «Сонные» трели обычны при согревании охлажденных яиц, двигательная активность эмбрионов в этом случае значительно снижена.
Непосредственно перед вылуплением эмбрионы «взрезают» скорлуповую оболочку яйца: этот процесс сопровождается специфическими «инструментальными» звуками, возникающими при трении яйцевого «зуба» о скорлупу. Интенсивность этих звуков чрезвычайно низка.
Выход птенцов из скорлупы сопровождается сигналами «вылупления». Их излучение вызывается болевыми ощущениями, поскольку в этот момент у птенцов происходит обрыв пупочного «стебелька». По спектрально-временным параметрам сигналы «вылупления» близки к звукам «дискомфорта»
Звуковая сигнализация на стадиях предвылупления у выводковых и полувыводковых птиц обеспечивает общение между эмбрионами в кладке, с одной стороны, и между эмбрионами и насиживающей птицей — с другой. Звуковое общение в этот период координирует поведение эмбрионов и приводит к установлению первичного акустического контакта с родителями, на основе которого после вылупления формируется стабильная связь взрослой птицы с выводком. Ритм сигналов «дискомфорта» у эмбрионов увеличивается при сходе (слете) птицы с гнезда. В этом случае они стимулируют возвращение насиживающей птицы. Магнитная запись звуков, издаваемых эмбрионами при естественной инкубации, позволила выявить некоторые особенности их звукового общения с насиживающей птицей. Так, излучение курицей-наседкой сигналов тревоги приводило к прекращению звуковой активности эмбрионов. Сход наседки с гнезда вызывал у эмбрионов спустя 5—8 мин интенсивные сигналы «дискомфорта», а возвращение птицы и ее призывные звуки активизировали «комфортную « сигнализацию. Воспроизведение для наседки звуков «дискомфорта» с помощью магнитофона приводило к тому, что он активно излучала призывные сигналы, перемещалась на гнездо постукивала клювом по скорлупе яиц. «Комфортные» сигналу эмбрионов не вызывали особых изменений в ее поведении.
Таким образом, формирование основных типов акустическихсигналов завершается перед вылуплением, что в последующем обеспечивает успешную акустическую ориентацию всего выводка Переход от акустического восприятия внешнего окружения, свойственного для эмбрионов, к восприятию комплексной афферентации после вывода сопровождается у птенцов дальнейшим развитием сигнализации. Появляются новые категории акустических сигналов, которые не наблюдались у эмбрионов: ориентировочны тревожные и тревожно-оборонительные. Вместе с этим происходит дальнейшее развитие сигналов «дискомфорта» и «комфорта».