 2015-10-13
2015-10-13 1119
1119Обширная группа имматуронатных видов (веслоногие, попугаи, стрижи, дятлы, воробьинообразные и т. д.) неоднородна по физиологической зрелости птенцов к моменту вылупления. У ряда групп (хищных птиц, сов, голенастых и некоторых других) птенцы вылупляются в различной степени опушенными, с закрытыми, полуоткрытыми или полностью открытыми глазами. Эту сборную группу условно относят к полуптенцовым или полувывлдковым.
На примере голенастых птиц установлено, то акустическая стимуляция эмбрионов приводит к акселерации эмбрионального развития и ускорению темпов вывода птенцов, однако очередность вылупления птенцов «повторяет» порядок откладки яиц. У птенцов, вылупившихся из «стимулированных» яиц полное раскрытие глаз происходит в среднем на сутки раньше по сравнению с птенцами, не подвергавшимися на стадиях предвылупления действию акустической стимуляции.
Акустически стимулы, применяемые до включения легочного дыхания, оказывают наиболее эффективное влияние на темп и синхронность вывода молодняка у куриных и пластинчатоклювых птиц. Привнесение стимуляции на более поздних стадиях эмбрионального развития (например, с момента выхода эмбриона в воздушную камеру яйца), хотя и ускоряет темп вылупления, но значительно слабее, чем при более ранней стимуляции. Причиной подобного явления может служить следующая закономерность эмбриогенеза выводковых птиц: стадиям предвылупления предшествует завершение роста эмбриональных тканей, после которого происходит некоторое снижение уровня метаболизма. Если акустическая стимуляция предшествует этому периоду, то он, как показывают результаты исследований, может быть укорочен или полностью элиминирован. Акселерация эмбрионального развития, таким образом, сопровождается возрастанием метаболических затрат. Ускорение метаболических процессов в эмбриогенезе птиц под действием акустической стимуляции предполагает ее влияние на центральные процессы, контролирующие метаболизм. Оно может заключаться в том, что активная «нагрузка» на анализаторные системы эмбрионов облегчает развитие и усиливает возбудимость нервных клеток, включенных в систему управления обменом, в частности регулирующих дыхательные процессы. Акустическая само- и взаимостимуляция эмбрионов (как впрочем и акустические внешние стимулы, близкие к видоспецифическим) существенно влияет на развитие периферического отдела слухового анализатора птиц, а также на развитие дендритной симметрии нейронах ламинарного ядра.
Серией экспериментальных исследований организации естественного поведения птенцов воробьиных птиц вскрыты механизмы ускоренного развития сенсорных и моторных компонентов основных функциональных систем (пищевод и оборонительной), полноценное функционирование которых определяет синхронное завершение гнездового периода. К главнейшим факторам синхронизации вылета из гнезда разновозрастных птенцов относят: (1) постоянное пребывание «младших» птенцов в сенсорнообогащенной среде, создаваемой вокализацией и перемещениями птенцов старшего возраста, (2) стабилизацию массы тела птенцов младшего возраста перед вылетом, в процессе которой у них отсутствует замедление роста («плато»), характерное для старших птенцов. Следует подчеркнуть, что подобное «замедление» роста, как уже отмечалось, свойственно для матуронатных птиц перед вылуплением, но этот период сниженного метаболизма может элиминироваться под действием акустической стимуляции. Как и в случае обязательных сроков акустической стимуляции эмбрионов выводковых птиц, необходимых для получения максимального эффекта ускоренного и синхронного вывода, для полноценного физического созревания в сокращенные сроки младшим птенцам у имматуронатных птиц необходим длительный контакт с птенцами старших возрастов.
К основным факторам, ускоряющим развитие птенцов, относят: повышенную двигательную активность, дополнительный комплекс тактильных и температурных воздействий, вызванный перемещением старших птенцов, и вокализацию, сопровождающую пищевую реакцию старших птенцов с первых дней жизни младших. Совокупность всех этих факторов, определяющая опережающее включение сенсорных и моторных компонентов функциональных систем гнездового периода птенцов, рассматривается как физически и социально обогащенная среда. Согласно концепции системогенеза, общие и частные закономерности онтогенеза функциональных систем находятся под определяющим действием экологических факторов, прежде всего это касается поведения.
Анализ акустической структуры сигналов и строения гортани. Многообразие морфологических типов нижней гортани в разных систематических группах птиц предполагает различие в уровне управления вокализацией у этих птиц.
Примитивная нижняя гортань представлена трахеей с преобладанием хрящевых колец, со слабой популунной складкой или без нее. Число мышц – 1-2 или значительно редуцированы. Можно предположить, что такие группы сохраняют «птенцовый» тип звуков без значительного нервного контроля за натяжением мускулатуры нижней гортани.
Нервный контроль за натяжением слаборазвитой сирингиальной мускулатуры у птенцов и взрослых неворобьиных птиц с относительно примитивным типом гортани сведен до минимума. Напротив, у воробьиных нервная регуляция имеет решающее значение как средство контроля за многомускульным управлением сиринкса, особенно при излучении сложных по формам частотной модуляции песен и позывов.
 У неворобьиных птиц со слаборазвитой мускулатурой нижней гортани определенную роль при звукоизлучении играет вытягивание шеи, ведущее к изменению положения трахеи и бронхов и о беспечивающее относительно стабильный уровень натяжения тимпанальных мембран. Хотя воробьиные птенцы имеют многомускульную нижнюю гортань, стабилизация контроля за натяжением тимпанальных мембран осуществляется в онтогенезе поэтапно,- например, у птенцов коньков, трясогузок и жаворонков, у которых это явление имеет место в 5—7-дневном возрасте. Это достигается на основе стабилизации дыхания «дозревания» всего комплекса голосовых мышц и включения ме ханизмов нервной регуляции их натяжения.
У неворобьиных птиц со слаборазвитой мускулатурой нижней гортани определенную роль при звукоизлучении играет вытягивание шеи, ведущее к изменению положения трахеи и бронхов и о беспечивающее относительно стабильный уровень натяжения тимпанальных мембран. Хотя воробьиные птенцы имеют многомускульную нижнюю гортань, стабилизация контроля за натяжением тимпанальных мембран осуществляется в онтогенезе поэтапно,- например, у птенцов коньков, трясогузок и жаворонков, у которых это явление имеет место в 5—7-дневном возрасте. Это достигается на основе стабилизации дыхания «дозревания» всего комплекса голосовых мышц и включения ме ханизмов нервной регуляции их натяжения.
Необходимо подчеркнуть, что увеличение скорости воздушного потока и его объема при генерации звуков на полном выдохе в условиях отсутствия или слабого нервного контроля за голосовой мускулатурой (например при излучении сигналов «дискомфорта» и пищевых сигналов в максимальной фазе пищевой peaции маленьких птенцов воробьиных птиц), обусловливает их широкополосную, амплитудно- или частотномодулированную структуру. Подобный тип звукоизлучения сохраняется у взрослых птиц в наиболее древней форме позывов - сигналах бедствия.
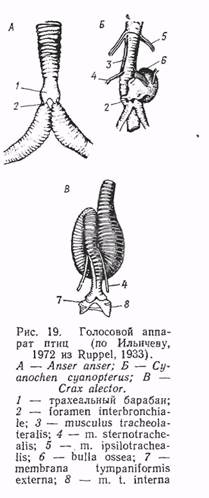
Нижняя гортань трахобронхиального типа, свойственная воробьиным, имеет сложную голосовую мускулатуру. Голосовые мышцы у воробьиных имеют максимальное развитие и их число достигает 7—9 пар, тогда как у более «примитивных» групп — гагар, поганок, веслоногих, голенастых, куриных, пластинчатоклювых и т. д.— их всего одна пара.
В простейшем случае механизм функционирования нижней гортани можно представить следующим образом: сокращение идущих вдоль трахеи грудинно-трахейных мышц прижимает трахею к бронхам и сближает голосовые (тимпанальные) мембраны, а козелок давит на выступ клавикулярного мешка, выпячивающего внутреннюю тимпанальную мембрану. Прохождение воздушной струи заставляет мембраны вибрировать. Таким образом производятся звуки в гортани птиц, имеющих слаборазвитую мускулатуру нижней гортани.
Значительно усложняется функционирование нижней гортани у воробьиных птиц. Грудинно-гиоидные мышцы, которые управляются подъязычным нервами, обеспечивают сложные и разнообразные движения элементов нижней гортани. особенно важна роль четырех пар трахеальных полуколец, пяти трахеальных колец наружных и внутрен-них голосовых мембран и шести пар этих мышц. Совместное действие этих элементов в момент производства звуков поворачивает четвертое бронхиальное полукольцо и приближает наружную голосо вую мембрану к внутренней. Поворот третьего бронхиального полукольца приближает переднюю часть внутренней голосовой мембраны к наружной мембране. Быстрое прохождение струи воздуха сближает мембраны и заставляет их вибрировать. Натяжение внутренней мембраны действием спинной ларинго-сирингиальной мышцы или задней бронхиальной влияет на частоту вибрации и размеры просвета бронхов и т. д.
Кроме перечисленных выше образований в звукоизлучении принимают участие, так или иначе влияя на спектрально-временные характеристики сигналов, трахеальные мешки (у эму), передний отдел пищевода (у африканского страуса и выпи), полость киля грудины (у журавлей), глоточные мешки (у дроф), пищевод (у голубей), клавикулярные и перивикулярные воздушные мешки (у многих видов воробьиных птиц). Эти резонаторы с мягкими стенками меняют свою форму и объем при действии мускулатуры, изменении положения головы, шеи, тела конечностей и т. п.
 Таким образом, производство акустических сигналов у птиц (особенно у воробьиных) связано со сложной системой морфологических образований, различных по структуре, происхождению и акустическому значению. Именно поэтому вопросы нервного контроля, точной координации являются одними из наиболее важных и наименее изученных в биоакустике птиц. Подход к решению этой задачи осложняется, прежде всего, тем, что в звукоизлучении у птиц участвуют мышцы различного центрального подчинения. Среди них выделяются непосредственно голосовая мускулатура, определяющая натяжение тимпанальных мембран и положение губ, мускулатура грудной клетки, определяющая выдыхание и прохождение воздушной струи через про' свет голосовых щелей в бронхах и трахее, а также мускулатура верхней гортани, регулирующая просвет гортанной щели, мускулатура шеи и головы, влияющая на расположение и длину трахеи, объем ротовой полости и т. д. Совместна работа всех этих образований достигает высокой степени согласованности, позволяя эффективно управлять звукоизлучением, и меняя в широких пределах частотные и амплитудные характеристики, а также временной «рисунок» сигнала.
Таким образом, производство акустических сигналов у птиц (особенно у воробьиных) связано со сложной системой морфологических образований, различных по структуре, происхождению и акустическому значению. Именно поэтому вопросы нервного контроля, точной координации являются одними из наиболее важных и наименее изученных в биоакустике птиц. Подход к решению этой задачи осложняется, прежде всего, тем, что в звукоизлучении у птиц участвуют мышцы различного центрального подчинения. Среди них выделяются непосредственно голосовая мускулатура, определяющая натяжение тимпанальных мембран и положение губ, мускулатура грудной клетки, определяющая выдыхание и прохождение воздушной струи через про' свет голосовых щелей в бронхах и трахее, а также мускулатура верхней гортани, регулирующая просвет гортанной щели, мускулатура шеи и головы, влияющая на расположение и длину трахеи, объем ротовой полости и т. д. Совместна работа всех этих образований достигает высокой степени согласованности, позволяя эффективно управлять звукоизлучением, и меняя в широких пределах частотные и амплитудные характеристики, а также временной «рисунок» сигнала.
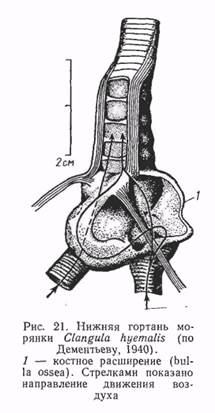
Следует подчеркнуть, что значительное влияние на процесс звукоизлучения у птиц оказывает трахея — резонатор широкого спектра действий. Длина трахеи у различных групп птиц варьирует в широких пределах: от 30 трахеальных колец — у мелких воробьиных до 400 — у журавлей и фламинго. Изменчива и форма трахеи. У куриных и некоторых воробьиных птиц она сужается в своей нижней части, у бескилевых и попугаев сжимается в дорсовентральном направлении. У самцов пластинчатоклювых и котинг (воробьиные) нижняя часть трахеи образует тонкостенные костные камеры разнообразной формы, достигающие значительных размеров. Половые различия в морфологии нижней гортани у пластинчатоклювых выражены уже на эмбриональных стадиях развития, а особенности механизмов звукоизлучения складываются в раннем онтогенезе н служат основой для различий в спектрально-временной структуре звуковых сигналов у самок и самцов сразу после вылупления.

