 2013-12-31
2013-12-31 785
785Окрашенные соединения типа пиррола.
Кроме птеридиновых пигментов, бактерии рода Pseudomonas синтезируют красные пигменты — антибиотики, принадлежащие к веществам типа пиррола: пиолутеорин, пирролнитрин, изопир-ролнитрин. Эти соединения были выделены из комплекса других окрашенных веществ в кристаллически чистом виде и установлена их химическая формула. Все эти вещества обладают антибиотической активностью против грамполо-жительных и грамотрицательных бактерий, дрожжей и грибов.
В работах последнего времени описан ряд штаммов бактерий рода Pseudomonas, которые, развиваясь на специфических синтетических средах, синтезируют внутриклеточные нерастворимые в воде пигменты желтого, розового и красно-оранжевого цвета. Использование современных методов исследования (хроматография, электрофорез и т. д.) позволило установить каротиноидную природу этих соединений.
Так, Ps. echinoides и его мутантные формы, а также Ps. methanica при развитии на синтетической среде синтезируют комплекс желто-оранжевых пигментов каротиноидного типа.
Большое количество пигментов, синтезируемых псевдомонадами, обладает фоточувствительностью, флуоресценцией и различной степенью растворимости. Биологическая роль и пути синтеза этой обширной группы соединений почти не изучены. Тщательное исследование пигментов, образуемых псевдомонадами, необходимо не только для уточнения систематики этого рода, но и для изучения физиологии отдельных видов и штаммов псевдомонад.
Род галобактериум (HALOBACTERIUM)
К этому роду относятся облигатные галофи-лы, способные развиваться в питательных растворах, содержащих не менее 12% соли (NaCl). Экстремальные галофилы выделены норвежским ученым X. Ларсеном из насыщенных растворов солей. При понижении концентрации солей в окружающей среде клеточная стенка бактерий разрушается. Палочковидные клетки превращаются в шаровидные сферопласты. При дальнейшем понижении концентрации соли клетки взрываются, так как внутриклеточное осмотическое давление оказывается очень высоким. Как было изучено, энзимы этих экстремальных галофилов активны лишь при высокой концентрации солей К и Na. Такие микроорганизмы, являясь гетеротрофами и используя энергию органических соединений, погибают в присутствии избытка пищи при поступлении воды в среду, понижающей концентрацию солей. Они не могут не быть галофилами. Являясь представителями своеобразной группы «высоленных форм» жизни, эти микробы обречены на существование в условиях соляной рапы. Любопытно отметить, что поиски этих форм были начаты с порчи рыбы и цветения соли в соляных испарительных бассейнах США (цветение соли отражает массовое развитие экстремальных галофильных бактерий, образующих розовый пигмент).
Род бделловибрио (BDELLOVIBRIO)
Исключительный интерес представляют недавно описанные немецким исследователем Г. Штольпом (1963) бактерии, паразитирующие внутри клеток других грамотрицательных бактерий. В пределах рода достоверно описан один вид — Bdellovibrio bacteriovorus. Бактерия представляет собой мелкую грамотрица-тельную, слегка изогнутую палочку. Размеры клетки: длина — 1 мкм, толщина — 0,3 мкм. Клетки снабжены одним полярно расположенным толстым жгутом (толщина 28 нм по сравнению с типичным жгутиком бактерий, толщиной 15—20 нм). Прикрепляясь к клетке-хозяину, паразит проникает внутрь клетки и развивается там. Именно все эти свойства побудили первооткрывателей дать этим бактериям приведенное выше название. Латинское слово «Bdello», означающее «пиявка», в сочетании с «vibrio» (вибрион) составило родовое название. «Vorus» — значит «пожирающий» — в сочетании с «bacterio» составило видовое название. В итоге эти, несомненно, интересные бактерии получили интригующее название «пиявко-вибрион, пожирающий бактерии».
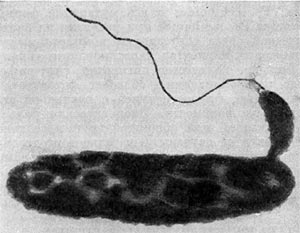
Рис. 22. Прикрепление паразитической бактерии Bdellovibrio к клетке хозяина — Pseudomonas (по Штольлу, 1963).
Немало хлопот доставляет этот организм исследователям ввиду его очень мелких размеров: он проходит через мелкопористые фильтры с диаметром пор, равным 0,45 мкм и даже 0,22 мкм, на которых задерживаются другие бактерии. Bdellovibrio bacteriovorus отличается от подобных ему бактерий прежде всего по тонкому строению жгутика. Он состоит из двух частей — сердцевины (толщина 13 нм) и чехла (7,5 нм), который покрывает жгутик. Чехол является продолжением клеточной стенки на жгутике. На противоположном полюсе клетка-паразит оснащена слизистой маской, лежащей сверху на складках, образованных клеточной стенкой. Этой частью паразит прикрепляется к поверхности клетки-хозяина (рис. 22). Здесь же можно часто видеть очень тонкую (45—55 А) короткую (0,8 мкм) нить, которая, как предполагают, подтягивает клетку-паразита к атакуемой клетке-хозяину. По тонкому строению клетки-паразита неотличимы от обычных грамотрицательных бактерий. Лишь мелкие размеры и способ добывания пищи выделяют их — они проникают внутрь клеток-хозяев и там развиваются (рис. 23 и 24).
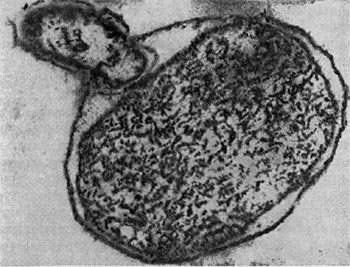
Рис. 23. Прохождение паразита внутрь клетки-хозяина. Ультратонкий срез (по Штольпу, 1963).
Клетки-паразиты не случайно сталкиваются с клетками-хозяевами, а активно ищут их. К слову, следует заметить, что паразиты перемещаются быстрее, чем многие другие клетки бактерий. Если поиск увенчался успехом, то можно под микроскопом увидеть удивительную картину преследования клетки-хозяина (какая - либо грамотрицательная бактерия) двумя-тремя клетками-паразитами. Преследуемая клетка активно сопротивляется, сбрасывая с помощью резких движений прикрепившихся паразитов. Иногда одну клетку-хозяина преследуют несколько клеток - паразитов. Такое разыскивание, преследование и, наконец,внедрение в клетку-хозяина протекает бурно и напоминает атаку (так и называется этот процесс в литературе). Прикрепление паразита начинается с вращательного движения вокруг своей оси, но может быть и шарнирным движением конца клетки-паразита на клеточной стенке хозяина. Скорость вращения достигает 100 об/мин. Прикрепившиеся клетки-паразиты очень прочно связаны с клеткой-хозяином. Их невозможно отделить ни встряхиванием, ни ультразвуком. При длительной обработке они гибнут вместе с хозяином, но не отделяются.
При вращении клетка-паразит пробуравливает клеточную стенку хозяина (см. рис. 23); паразит, проникнув внутрь клетки-хозяина, оттесняет протопласт и располагается между клеточной стенкой и цитоплазматической мембраной, где он многократно размножается. Клетка-хозяин похожа на «мешок», набитый паразитами (20—50 клеток-паразитов в одной клетке-хозяине; рис. 25). Эти интересные наблюдения проведены советскими исследователями Е. Н. Мишустиным и Э. С. Никитиной.
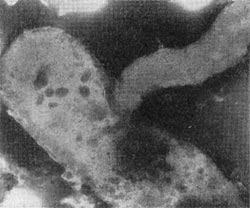
Рис. 24. Атака паразитической бактерией клетки-хозяина. Увел, х 35000.
В настоящее время известно довольно много бактерий, различающихся морфологически, которых относят к роду Bdellovibrio, причем наряду с мелкими вибрионоподобными клетками встречены довольно крупные палочки. Это не должно удивлять, так как в процессе проникновения в клетку через сравнительно маленькое отверстие клетка-паразит сужается и как бы продавливается через отверстие, «просачиваясь» внутрь клетки-хозяина.
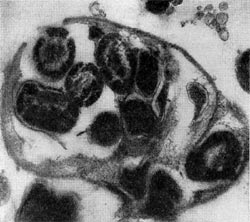
Рис. 25. Клетка бактерии-хозяина (Tuberoidobacter sp.), заполненная размножившимися паразитами. Электронная микрофотография. Увел, х 40000.
Процессы атаки, внедрения паразита внутрь клеток-хозяев протекают очень быстро. Спустя 2—3 мин после атаки клетки-паразиты внедряются в клетки-хозяев. По истечении 10— 60 мин они полностью проникают в хозяев, а через 3—5 ч, завершив разрушение атакованных ими клеток-хозяев, паразиты освобождаются из разрушенных клеток. Инфицированная паразитом клетка уже вскоре после атаки подвергается большим структурным изменениям, что связано в первую очередь с нарушением целостности и прочности клеточной стенки. Изменяются контуры клетки — возникают шаровидные сферопласты. Вслед за стенкой идет разрушение содержимого клеток.
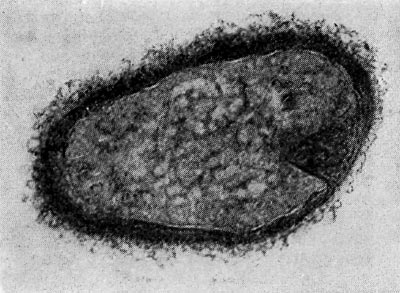
Рис. 26. Цисты (форма покоя) паразитических бактерий Bdellovibrio. Увел. X 50000.
Наряду с паразитическими формами Bdellovibrio возникают сапрофитные, образующие желтый пигмент. Е. Н. Мишустину и Э. С. Никитиной удалось доказать, что часть паразитов, выбравшись из обломков разрушенной ими клетки-хозяина, переходит не к сапрофитному способу питания, а в состояние покоя, образуя цисты. Это овальные, почти круглые клетки с уплотненной ядерной зоной и толстой оболочкой. Размеры их необычайно малы, и они оставались не замеченными многими микробиологами (рис. 26).
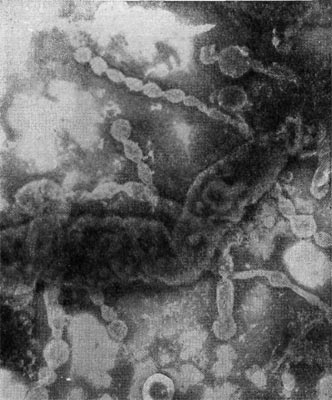
Рис. 27. Разрушенная клетка неспороносной палочки. Видны выброшенные из клетки цепочки внутриклеточных мембран (мезосом). Увел. X 25000.
Особый интерес представляет конечный результат атаки и развития паразитов в клетках других бактерий — их разрушенные (лизиро-ванные) остатки. Такие остатки (детрит) часто обнаруживаются в субстратах, где живут микробы. Они остаются после отмирания стареющих клеток микробных популяций, особей, лизированных фагами, и др. Все указанное относится к эндогенному типу лизиса, когда действующий фактор (в случае отмирания старых клеток сказывается разрушение клетки изнутри ферментами) находится внутри клетки и его действие направлено от центра к поверхности клетки. Bdellovibrio атакует клетку сверху, нарушая целостность клеточной стенки и лишь затем проникая внутрь. Клеточная стенка хозяина разрушается под влиянием ферментов паразита, обнажая ленты мембранных структур (рис. 27). Атакованные паразитами клетки с нарушенной стенкой часто образуют вздутые за счет внутреннего осмотического давления клетки (сферопласты), из которых вышвыриваются наружу цепочки мембран -внутренние структуры (интрацеллюлярные мембраны), на которых ориентированы ферменты, обеспечивающие важнейшие жизненные функции клетки. Эти остатки клеток-хозяев, убитых бактериями-паразитами, еще долгое время остаются в среде, лишь постепенно разрушаясь под влиянием уже других, обычных сапрофитов — «мусорщиков», но все еще остаются на поле битвы следы веществ — индикаторов бурных процессов гибели клеток — составные части клеточных стенок бактерий: мурамовые кислоты, составляющие мешок, покрывающий клетки.
Паразитические бактерии Bdellovibrio широко распространены в природе. Они встречаются в воде рек, озер, морей, в почвах разных географических зон в сравнительно небольших количествах, как бы выжидая момента атаки в случае появления большого количества клеток других бактерий, в первую очередь больных. В этом случае активизируются не только клетки паразита, но и его формы покоя — цисты, которые быстро прорастают. Bdellovibrio выполняют роль волков в микробном сообществе, выедая не всю, а часть популяций клеток (никогда не наблюдалось полного уничтожения хозяев). Объектами атаки (хозяевами) паразитов являются практически все бактерии, но в первую очередь грамотрицатель-ные, в том числе возбудители опасных эпидемических болезней, например холеры. Использование Bdellovibrio (предварительно накопленного в лабораторных условиях) для обеззараживания воды в водоемах уже нашло применение. Характер взаимоотношений Bdellovibrio-пaразита с другими бактериями-хозяевами является ярким примером широко распространенного в живом мире нашей планеты одного из трех способов добывания пищи. Два других — сапрофитный и хищный способ добывания пищи. Тем не менее между этими способами нет больших различий. Сапрофиты питаются мертвыми микроорганизмами, хищники убивают сами или пользуются услугами других «убийц», а паразиты поедают своих хозяев постепенно, совсем не стремясь их убить, по крайней мере немедленно. Таким образом, все живые существа могут быть разделены на «пожираемых» и «пожирающих», и исследование явления паразитизма на любом уровне представляет общебиологический интерес.








