 2013-12-31
2013-12-31 834
834Существование у растений приобретённого иммунитета, как у теплокровных животных и человека, до недавнего времени ставилось под сомнение на том, что у них отсутствуют нервная система и кровообращение. В настоящее время факт существования приобретённого иммунитета уже не вызывает никаких сомнений. Сведения о наличии цитоплазматической связи между клетками при помощи плазмодием, проходящих через поры клеточных стенок, оказались весьма полезными в изучении этого явления. Через плазмодиями раздражение определённого участка растительной ткани распространяется практически по всем клеткам растения. Воздействие инфекции проявляется не только в клетках, непосредственно контактирующих с патогенном, но и в удалённых от места инфекции. Так, при заражении плодов цитрусовых возбудителем голубой плесени (Penicillium italicum) интенсивность дыхания кожуры возрастает не только в очаге инфекции, но и в неинфицированных тканях, включая расположенные на противоположной стороне плода.
В зависимости от причин, вызвавших приобретенный иммунитет, его разделяют на инфекционный и неинфекционный. Инфекционный приобретенный иммунитет возникает в результате перенесенного растением заболевания, неинфекционный приобретенными или индуцированный, иммунитет — под влиянием внешних факторов, не приводящим к изменению генома растения. Факторы, воздействие которых на семена или растения приводит к повышению устойчивости растений, называются индукторами. Неинфекционный приобретенный иммунитет играет важную роль в практике сельского хозяйства и рассматривается как один из способов защиты от болезней.
Приобретенный иммунитет может иметь локальный или системный характер.
Локальный иммунитет проявляется вблизи места воздействия индуктора, системный — в участках, удаленных от этого места, например в других или новых листьях. Приобретенный иммунитет, как правило, неспецифичен.
Индукторы приобретенного иммунитета делят на биотические и абиотические. К первым относят грибы, бактерии, вирусы или продуцируемые ими метаболиты, ко вторым — химические вещества (биорегуляторы) или их смеси и физические воздействия (например, облучения, температуры, магнитного поля, ультразвуковых колебаний и т.д.).
Индуцирование биотическими средствами проводят путем инокуляции авирулентными (непатогенными) расами, инактивированными патогенами и продуктами их метаболизма.
Индуцирование приобретенного иммунитета биотическими средствами, называемое иммунизацией, сходно с вакцинацией, широко используемой в медицине и ветеринарии.
Изучение явления иммунизации имеет длительную историю, однако в отличие от вакцинации она не нашла пока широкого практического применения в защите растений.
Вакцинация растений как способ биологической иммунизации находит применение в практике сельского хозяйства, особенно в создании приобретенного иммунитета к вирусным болезням у овощных культур — представителей семейств пасленовых (томата) и тыквенных. Заражение рассады этих культур слабыми штаммами некоторых вирусов приводит к появлению у растений иммунитета, благодаря которому они становятся устойчивыми к сильнопатогенным штаммам тех же вирусов. Этот прием получил название перекрестной защиты или интерференции.
Повышение устойчивости растений к патогену можно вызвать путем предварительного инфицирования их непатогенным изолятом. Так, клубни картофеля, предварительно инфицированные авирулентным штаммом Ph. infestans, приобретают устойчивость к вирулентному изоляту этого гриба. В качестве вакцины могут быть использованы убитые патогены. Так, введение в ткани листьев табака убитой культуры Pseudomonas tabacum индуцирует устойчивость к этому патогену. Установлено, что в результате вакцинации в клетках растения появляются защитные вещества, например фитоалексины, изменяется активность ряда ферментов и появляются новые. При приобретенном иммунитете в отличие от врожденного защитные механизмы возникают в ответ на вакцинацию и не передаются по наследству.
Рострегулирующие бактерии, обитающие в ризосфере, могут защищать растения от грибных, бактериальных и вирусных болезней, индуцируя их системную устойчивость. Ризосферные микроорганизмы удобны для применения в целях индуцирования устойчивости растений к болезням. Их можно нанести на семена перед посевом или на корневую систему рассады перед посадкой путем обработки суспензией бактерий и грибов. При этом ризообитаю-щие непатогенные микроорганизмы могут выступать конкурентами для патогенов, колонизируя ткани растений, либо выделять метаболиты, включая те, которые обусловливают защитные реакции растений против патогенов. Индукционную активность на ряде культур проявляет препарат триходермин, созданный на основе гриба Trichoderma lignomm.
К настоящему времени выявлено большое число синтетических и природных соединений, индуцирующих устойчивость растений к болезням.
Аминокислоты и их аналоги. В культуральной жидкости биологических индукторов устойчивости растений к болезням обнаружены свободные природные аминокислоты. Наибольшей индукционной активностью обладает фракция, содержащая в основном глутаминовую кислоту, пролин, лейцин и аланин. Эта фракция снижала на 54 % развитие грибов рода Fusarium на проростках пшеницы (Сокирко, Вовчук, Гаврилов, 1989).
Производные карбоновых кислот. Среди производных карбо-новых кислот выявлены вещества, оказывающие влияние на устойчивость растений к болезням. Среди них следует отметить салициловую кислоту, которая специфически связывается в растениях с каталазой и ингибирует активность этого фермента, что сопровождается накоплением пероксида водорода— одного из основных защитных веществ растений от патогенов. Она также оказывает влияние на уровень фитогормонов (индолилуксусной и абсцизовой кислот, а также этилена) в растениях. Индуцирующей активностью обладают и ненасыщенные высшие карбоновые кислоты, такие, как арахидоновая, линоленовая, линолевая, жасмо-новая, абсцизовая и полиакриловая. Арахидоновая кислота входит в состав диглицеридов, принимающих участие в установлении межклеточных контактов и передающих информацию с поверхности клетки. Арахидоновая кислота стимулирует в растениях защитные реакции и индуцирует накопление фитоалексинов. В малых концентрациях она повышает устойчивость растений картофеля к фитофторозу. В клубнях,' обработанных арахидоновой кислотой, устойчивость к этому заболеванию развивается на вторые сутки и сохраняется от нескольких недель до" 2...3 мес. Она носит системный характер.
Смесевые препараты. Один из таких препаратов — хитозан, состоящий из глюкозаминов, индуцирует устойчивость растений риса к пирикуляриозу и ризоктониозу. К препаратам комбинационного действия следует отнести четвертичные соли аммония, содержащие в своем составе анион аминобензойной кислоты и катион четвертичного аммония с гидроксиалкильной группой. Предпосевная обработка семян зерновых, овощных и некоторых других культур четвертичными солями аммония повышает устойчивость растений к корневым гнилям, уменьшает агрессивность фитопатогенов и улучшает показатели структурных элементов урожайности на 20...25 % (Шкаликов, Хохлов, 1998).
Повышение устойчивости растений к болезням наблюдалось также после предпосевной обработки семян смесями микроэлементов (Кулешова, 1990). О роли микроэлементов в жизнедеятельности растений свидетельствует тот факт, что существует целая группа болезней неинфекционного характера, которые возникают только в случае отсутствия или недостатка в почве каких-либо микроэлементов. Так, при недостатке в почве бора возникает гниль сердечка сахарной свеклы, развивается бактериоз льна, вызываемый Bacillus macerans. Неинфекционный хлороз плодовых деревьев проявляется при недостатке в почве железа. Возможность использования удобрений для повышения устойчивости растений к болезням была обоснована Т.Д. Страховым (1922). Он впервые установил, что, регулируя режим питания растений, можно изменить их устойчивость к тому или иному заболеванию. Он показал также, что удобрения могут повышать устойчивость злаковых культур к головневым болезням. Сходные данные были получены в многочисленных исследованиях различных сочетаний растение — паразит. Механизм действия удобрений на устойчивость растений многообразен. Они оказывают существенное влияние на ход биохимических и физиологических процессов, а следовательно, и на обмен веществ растения, изменения в котором могут сказываться на взаимоотношениях между растением и паразитом, в основе которых лежит приуроченность патогена к определенному типу обмена веществ растения. Чем выше паразитическая специализация патогена, тем теснее взаимосвязь между обменом веществ паразита и растения-хозяина. Так, при высоком содержании азота в почве растения обычно более восприимчивы к болезням. Многие облигатные паразиты (например, возбудители мучнистых рос, ржавчинных болезней и т. д.) лучше развиваются на растениях с мощной вегетативной массой. Калийные и фосфорные удобрения способствуют повышению устойчивости растений. Однако сделать общие выводы о влиянии удобрений на поражаемость растений достаточно сложно, поскольку они могут ускорять или замедлять рост и развитие растений, что, в свою очередь, влияет на устойчивость к тому или иному патогену. Например, азотные удобрения в большинстве случаев снижают устойчивость растений к болезням, однако в отношении головневых грибов наблюдается обратная связь. Возможно, более быстрый рост растений, наблюдаемый при внесении азотных удобрений, помогает им быстрее пройти восприимчивую фазу развития. Таким образом, научно обоснованное применение как макро-, так и микроудобрений повышает способность растений противостоять болезни.
Защитные реакции растений изменяются при воздействии на сложившиеся между ним и паразитом взаимоотношения. Так, с помощью приемов, ускоряющих прохождение отдельных фаз развития хозяина, можно сократить возможный период его заражения. Например, обрезка деревьев, кустарников, подкашивание трав (клевера, люцерны) повышает устойчивость к таким приуроченным к периоду старения болезням, как антракноз, серая гниль и др.
Исходя из разнокачественности семян используют специальные методы их отбора, позволяющие выращивать растения с повышенной устойчивостью. Известно, что из здоровых полновесных семян развиваются растения, менее восприимчивые к болезням. Поэтому посев семян, отсортированных по удельной массе, рекомендован как один из методов снижения поражаемости пшеницы фузариозом или кукурузы пузырчатой головней.
В практике сельского хозяйства известно немало подобных приемов, способствующих повышению устойчивости. В большинстве своем они основаны на знании закономерностей проявления иммунитета, что дает возможность направленного создания условий, благоприятствующих проявлению у растений защитных свойств.
3. ТИПЫ ПАРАЗИТИЗМА У МИКРООРГАНИЗМОВ
Паразиты заселяют растительную ткань только в том случае, если она содержит необходимые им питательные вещества. Таким образом, паразитизм основан на пищевых взаимосвязях между патогеном и растением-хозяином. Что касается эволюции паразитизма, то некоторые исследователи полагают, что паразиты растений происходят от форм, ведущих сапротрофный образ жизни на стареющих растительных тканях, т. е. эволюция шла в направлении от облигатных сапротрофов к облигатным паразитам.
Для облигатных сапротрофов источником питательных веществ служат только мертвые органические субстраты. Факультативными паразитами называются организмы, которые обычно живут как сапротрофы, но при определенных условиях могут нападать на живые ткани, в частности на старые, ослабленные и поврежденные или на запасающие органы. К таким паразитам относятся, например, Botrytis cinerea, Rhyzopus nigricans и др. Такие пластичные патогены часто имеют широкий круг хозяев, отличаются слабой специализацией и хорошо растут на искусственных питательных средах. Факультативные сапротрофы— это организмы, которые обычно ведут паразитический образ жизни, но могут существовать и как сапротрофы. Примером таких организмов могут служить Venturia inaequalis, Phytophthora infestans и др. Они имеют более узкий круг растений-хозяев и более выраженную специализацию, в культуре растут медленно. Экологически облигатными паразитами принято считать такие патогены, которые существуют в природе как паразиты, но их можно выращивать и в культуре на искусственных питательных средах. К таким паразитам относят многие виды Ustilaginales. Облигатными паразитами называются организмы, например Erysiphales, Uredinales, Peronosporaceae, Albuginaceae, живущие только как паразиты.
Патогенность некоторых паразитов ослабевает при выращивании в аксеничной культуре. По-видимому, это объясняется тем, что в таких условиях определенные преимущества приобретают сапротрофные штаммы. Возможно также, что в основе этого явления лежит адаптация. Однако часто патогенность удается восстановить путем пассажей через восприимчивые растения.
Паразиты по-разному извлекают питательные вещества из растений-хозяев. В связи с этим их можно разделить на две группы — биотрофов и некротрофов. Биотрофы извлекают необходимые им питательные вещества из живых клеток растений-хозяев, н е кротрофы — из мертвых. Как правило, облигатные паразиты не убивают восприимчивые клетки, в которые проникают: им это не выгодно, поскольку только живые клетки могут быть источником питательных веществ.
Способность патогенных микроорганизмов паразитировать на растениях обусловлена специфическими особенностями их обмена веществ, позволяющими им преодолевать защитные механизмы растений-хозяев и использовать содержимое их клеток в качестве питательного субстрата. Механизм использования разнообразных питательных веществ варьирует у различных фито-патогенов в весьма широких пределах. Он обусловлен приспособленностью этих паразитов к тому или иному виду растений, а также к химическому составу их клеток и анатомо-морфологическим особенностям тканей. Так, факультативным паразитам свойственна способность использовать широкий набор питательных веществ. Они образуют множество разнообразных ферментов, которые расщепляют полимерные пластические вещества растений, доводя их. до усвояемой формы. У некоторых патогенов (например, Poliporus) синтез экзоферментов (внеклеточных ферментов) значительно превышает синтез внутриклеточных. Это означает, что большая часть ферментов, образующихся в гифах факультативных паразитов, выделяется в окружающую среду.
В литературе накоплено множество данных о вооруженности ферментами грибов — факультативных паразитов и сапротрофов. Среди экстрацеллюлярных ферментов названных грибов имеются разнообразные карбогидразы, осуществляющие гидролиз различных ди- и полисахаридов (инулина, крахмала и др.), гемицеллю-лозы, клетчатки и т. п. Помимо выделения ферментов в окружающую среду микроорганизмы могут воздействовать на питающий их субстрат путем контакта. Установлено, что подобное действие зависит от ферментов, локализованных на поверхности клеток. Активность и качественный состав выделяемых ферментов в онтогенезе грибов меняется. С возрастом может меняться и активность одного и того же фермента. Показано, что низкая ферментативная активность приурочена к моменту прорастания спор. По мере развития гриба набор ферментов в его тканях расширяется и возрастает их активность. В стадии спорообразования она резко снижается.
Сравнительное изучение биохимических свойств факультативных и облигатных паразитов позволило установить, что прорастающие споры облигатного паразита менее богаты ферментами. Наиболее мощные протеолитические ферменты и амилазы у облигатных паразитов либо отсутствуют, либо обладают незначительной активностью. Поэтому внедрение в ткани растения-хозяина облигатных паразитов не вызывает резких нарушений в его обмене веществ.
Гибель клеток растения-хозяина обусловлена действием токсических для высшего растения веществ. Выявлены значительные различия в количестве и составе биологически активных веществ, синтезируемых фитопатогенами и они тем существеннее, чем четче выражены у патогенна паразитические свойства. Факультативные паразиты обладают способностью к образованию высокотоксичных веществ, вызывающих гибель тканей хозяина и создающих возможность успешного развития и распространения микроорганизмов. В большинстве случаев способность образовывать токсические вещества является признаком, выработавшимся и закрепившемся в процессе эволюционного развития и определяющим собой характер взаимоотношений паразита и растения-хозяина.
4. ГЕНЕТИКА, БИОХИМИЯ И МОЛЕКУЛЯРНАЯ БИОЛОГИЯ ИММУНИТЕТА РАСТЕНИЙ
4.1 ПОЛЕВАЯ И ИСТИННАЯ УСТОЙЧИВОСТЬ
Реакция растения на заражение инфекционными объектами находится под значительно более сложным генетическим контролем, чем другие хозяйственно ценные признаки, такие, как количество и качество продукции, срок созревания и т. п. Это обусловлено тем, что на степень пораженное™ растения оказывают влияние различные морфологические, анатомические, физиологические и биохимические факторы. Например, сорта пшеницы, имеющие высокую скорость прорастания и прохождения ранних фаз роста, слабо поражаются твердой головней, а такие же сорта свеклы и хлопчатника устойчивы к корневым гнилям, вызванным комплексом почвообитающих грибов, лишь потому, что указанные паразиты способны заразить только молодые проростки, не приобретшие еще плотных покровных тканей. Сорта картофеля с рыхлой, раскидистой ботвой слабее, чем сорта с плотной ботвой, поражаются фитофторозом в связи с тем, что влага после дождя или ночи с обильным образованием росы на листьях первых сортов лучше проветривается и быстрее высыхает, так что зооспоры не успевают освободиться из зооспорангиев, инцистироваться, прорасти и внедриться в ткань листа. Виды барбариса, имеющие толстую кутикулу и наружную стенку эпидермиса, более устойчивы к возбудителю стеблевой ржавчины злаков Puccinia graminis, чем виды с тонкими покровами, поскольку проростки монокарио-тических базидиоспор внедряются в лист непосредственно через эпидермис. В то же время устойчивость злаков к этой болезни никак не связана с толщиной кутикулы, так как проростки дикарио-тичных эцио- и урединиоспор проникают через устьица. Сорта злаков с пониклыми листьями менее поражаются возбудителями листовых пятнистостей, чем сорта с листьями, расположенными под острым углом к стеблю, лишь по той причине, что инфекционные капли скатываются с листьев на грунт. Рожь сильно поражается спорыньей, а пшеница очень слабо в связи с разными стратегиями размножения: у ветроопыляемой ржи цветки долго остаются открытыми для заноса в них спор, а самоопыляемая пшеница имеет закрытое цветение.
Все эти и многие другие примеры устойчивости можно обнаружить при полевой оценке сортов, но они могут не проявляться при искусственном заражении в лаборатории. Поэтому перечисленные примеры устойчивости объединяют общим понятием полевая устойчивость. Как видно, полевой устойчивостью управляют различные гены, и при отборе на её повышение часто используют косвенные признаки (скорость прохождения отдельных фаз онтогенеза, толщина кутикулы, габитус куста и т.д.), а не прямое заражение.
Альтернативная полевой – истинная устойчивость, которая проявляется не только в поле, но и при заражении отдельных от растения листьев в условиях, способствующих развитию болезни.
4.1.1. БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ИСТИННОЙ УСТОЙЧИВОСТИ
Если в основе полевой устойчивости лежат анатомоморфоло-гические и онтогенетические особенности растений, то истинная устойчивость обусловлена главным образом накоплением токсических для паразита продуктов. Растение продуцирует огромное число разнообразных веществ, несущих защитные функции. Эта вещества различаются по химическому составу (от простейшей органической кислоты — щавелевой до высокомолекулярных белков), мишеням (защищают от грибов, бактерий, вирусов, насекомых), механизмам действия (убивают патогенов, инактивируют орудия их агрессии — токсины, ферменты и др.), способам возникновения (присутствуют в интактных клетках, индуцируются заражением). Здесь будут рассмотрены особенности строения и механизма действия лишь некоторых представителей двух групп антибиотических веществ растений — низкомолекулярных соединений (вторичных метаболитов) и белков.
НИЗКОМОЛЕКУЛЯРНЫЕ ЗАЩИТНЫЕ ВЕЩЕСТВА. Их разделяют на два класса, названные фитоантиципины и фитоалексины. Первые представляют собой низкомолекулярные антимикробные соединения, присутствующие в растении до заражения или продуцируемые после механического повреждения клетки и заражения из предшественников. Вторые — низкомолекулярные антимикробные соединения, которые синтезируются и аккумулируются в растении после взаимодействия с паразитом.
Фитоантиципины. История изучения фитоантиципинов началась после того, как в 1928 г. зоолог-эмбриолог Б. П. Токин обнаружил, что летучие соединения, образующиеся в результате растирания лука и чеснока, убивают простейших животных (инфузорий и др.). Эти летучие продукты Токин назвал фитонцидами. Название было не очень удачным, ибо фитон по-гречески означает растение, а цидум по-латыни — убивать, то есть этот термин — гибрид из разных языков, что не положено; к тому же дословный его перевод — убивающие растения, хотя эти вещества продуцируются растениями, а убивают микроорганизмы. Тем не менее в отечественной фитопатологии этот термин прижился и получил широкое распространение, причем фитонцидами стали называть не только летучие, но и все остальные антимикробные соединения. Известный фитопатолог Д. Д. Ведеревский и его ученики проделали огромную работу по изучению влияния экстрактов растений на различные фитопатогенные объекты. К сожалению, информационная ценность этих исследований оказалась довольно низкой. Дело в том, что живая клетка представляет собой систему покрытых мембранами пузырьков (компартментов). Пока клетка жива, вещества, находящиеся в разных пузырьках, не взаимодействуют друг с другом, но если ткань мацерировать, то клетка окажется поврежденной и вещества из разных компартментов (вакуолей, лизосом, пероксисом и др.) придут в контакт друг с другом и в результате химических реакций возникнут соединения, часто токсичные, которых в неповрежденных клетках не было. Например, корень хрена сам по себе не вызывает никаких неприятных ощущений, но если его натирать, то образуются летучие соединения, токсичные для слизистых оболочек.
Большинство токсических веществ находится в живой клетке в форме гликозидов, то есть в соединении с одним или несколькими остатками Сахаров. Гликозиды менее токсичны, чем отделенные от остатков Сахаров агликоны, и поэтому неопасны для самой клетки; они водорастворимы и поэтому являются транспортной формой токсических веществ. Обычно они находятся в вакуолях и отделены тонопластом от других компонентов клетки, что также защищает ее от повреждающего действия. При нарушении целостности клеточных мембран содержимое вакуолей выливается в клетку и гликозиды входят в контакт с ферментами г л и к о -зидазами, находящимися в других мембранных пузырьках лизосомах или ковалентно связанными с полисахаридами клеточной стенки. Гликозидазы отщепляют сахара от гликозидов и освобождают высокотоксичные агликоны, убивающие клетку и находящихся в ней паразитов.
Классификация гликозидов основана на химическом строении агликонов.
Фенолы и фенольные гликозиды. Фенолами называют циклические соединения, содержащие ароматическое кольцо с фенильным гидроксилом или его замещенными радикалами (кислотным, альдегидным и др.). Многие растительные фенолы помимо фенила (С6) содержат пропановый радикал (С4) и названы фенилпропано-идами. Фенолы, с одной стороны, нормальные метаболиты растений, так как входят в состав регуляторов роста, пигментов, структурных элементов клеточной стенки (лигнина), а с другой — это стрессовые метаболиты, синтез которых резко возрастает при поранении или при заражении. Они образуются в результате дезами-нирования аминокислоты фенилаланина с помощью фермента фенилаланин-аммиаклиазы (ФАЛ), активность которой обычно возрастает при стрессорных воздействиях (заражении, механическом повреждении, облучении), почему ФАЛ называют стрессовым ферментом.
Основные механизмы антимикробного действия фенолов следующие.
• Продукты окисления фенолов полифенолоксидазами (ПФО) хиноны обладают высокой реакционной способностью. Они могут инактивировать белки вследствие формирования замещенных продуктов с SH-, NH2-rpynnaMH белков. Как сильные окислители, хиноны акцептируют электроны от многих соединений.
• Фенолы влияют на трансмембранный транспорт веществ в клетке. Некоторые замещенные фенолы служат проводниками протонов в мембранах и вследствие этого действуют как разобщающие агенты.
• Промежуточные соединения биосинтеза фенолов могут влиять на метаболизм фитогормонов. Являясь ингибиторами фермента ИУК-оксидазы, многие фенолы способствуют накоплению фитогормона индолилуксусной кислоты (ИУК), а гормональный статус растения оказывает существенное влияние на взаимоотношения хозяина и паразита.
• Фенолы принимают активное участие в создании химических барьеров. Продукты окисления фенольных спиртов (кумарилово-го, кониферилового, синапилового), конденсированные в результате свободнорадикальных реакций, образуют аморфный трехмерный полимер лигнин, откладывающийся в клеточных стенках (обычно во вторичных стенках и стенках мертвых клеток). Лигнин ковалентно связывается с полисахаридами клеточной стенки и защищает их от атаки грибными ферментами деполимеразами, препятствует диффузии в клетки грибных метаболитов, обладает прямой токсичностью для микроорганизмов, может лигнифициро-вать (ограничивать в росте) гифы грибов, проникшие в клетки растений.
Важную роль в образовании лигнина играет другой стрессовый фермент — пероксидаза, активность которого, как и активность ФАЛ, возрастает при механическом повреждении тканей и при заражении. Пероксидаза осуществляет полимеризацию коричных спиртов в лигнин, формирует прочные сшивки между целлюлозой, пектином, гликопротеинами клеточных стенок и лигнином, участвует в заживлении ран — отложении водонепроницаемого барьера из полимеризованных алифатических и ароматических соединений (суберинизации ткани), участвует в катаболизме (разложении) ауксинов. В растительной клетке пероксидаза локализована в разных компартментах и существует в нескольких изофор-мах.
• Катионные (рН 8,1..Л 1). Катализируют образование перокси-да водорода из НАДН и воды. Локализованы в центральной вакуоли. Обладают активностью ИУК-оксидазы и регулируют уровень ауксинов в клетке.
• Слабоанионные (рН 4,5...6,5). Локализованы в клеточных стенках. Умеренно активные лигнификаторы. Осуществляют су-беринизацию ран.
• Анионные (рН 3,5...4,0). Локализованы в клеточных стенках. Осуществляют лигнификацию и сшивки полисахаридов клеточных стенок. Их активность регулируется уровнем ауксинов.
Фенолы находятся в клетках растения в свободном виде или в форме фенольных гликозидов. Например, сорта лука с окрашенными (желтыми и красными) чешуями устойчивы к заражению многими грибами (из родов Aspergillus, Alternaria и др.). Их окраска обусловлена фенольным гликозидом флавоновым пигментом кверцитрином, находящимся в вакуолях. Как известно, кроющие чешуи лука состоят из мертвых клеток. При их отмирании происходит разрушение внутриклеточных мембран, вследствие чего от фермента гликозидазы отщепляется фенольный агликон кверцитин, который под действием другого фермента полифено-локсидазы (ПФО) расщепляется с образованием низкомолекулярных водорастворимых монофенолов — протокатеховой кислоты и ортокатехола (рис.8). Эти фенолы обладают высокой фунгитоксичностью и, диффундируя в инфекционную каплю, убивают споры грибов.
В листьях яблони содержится фенольный гликозид ф л о р и д -зин, который при механическом повреждении листьев или при заражении возбудителем парши Venturia inaequalis гидролизуется ферментом Р-гликозидазой, образуя фенол флоретини глюкозу. Флоретин окисляется фенолоксидазой с образованием высокотоксичного, но нестабильного хинона. Обработка листьев перед заражением химическими соединениями — ингибиторами гликозидазы и ПФО — превращает устойчивые сорта в восприимчивые к болезни.
Терпеноиды и терпеноидные гликозиды. К терпеноидам относят соединения, синтезируемые из уксусной кислоты путем конденсации ее молекул. Первый продукт конденсации — изопрен — содержит пять атомов углерода. В построении терпеноидов участвуют две группы ферментов — полиме-разы, соединяющие молекулы изопрена в цепочку, и циклазы, осуществляющие преобразование цепочек в замкнутые кольца. При конденсации двух молекул изопрена образуются соединения, названные монотерпенами (С10). Это низкомолекулярные летучие вещества, которые часто придают
 |
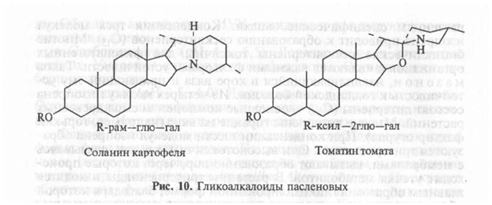
растениям специфические запахи. Конденсация трех молекул изопрена приводит к образованию сесквитерпенов (С 15). Многие бициклические сесквитерпены токсичны для фитопатогенных организмов и являются важными факторами устойчивости. Таков м а з о н о н, накапливающийся в коре вяза и придающий ему устойчивость к голландской болезни. Из четырех молекул изопрена состоят дитерпены (С20) — основные компоненты смол древесных растений. Многие дитерпены горькие на вкус (например, горький фактор огурца). При конденсации шести молекул изопрена образуются тритерпеноиды. Они высокотоксичны, так как, связываясь с мембранами, вызывают образование пор, через которые происходит утечка метаболитов. В растении тритерпеноиды находятся главным образом в гликозилированной форме, благодаря которой образуют мылящиеся в воде растворы: именно поэтому их называют сапонинами (сапо по-латыни — мыло). Сапонины обладают выраженным антигрибным действием. Например, сапонин цикламена цикламин подавляет рост грибов в концентрациях ниже 200 мкг/мл. Сапонины представляют собой токсические начала таких ядовитых растений, как наперстянка, ландыш, куколь и др. (встречаются у представителей более 70 семейств растений). Агли-коном сапонинов у одних растений может быть пентацикличес-кий тритерпеноид (рис. 9), у других — тетрациклический стероид. У пасленовых и некоторых других растений к стероидному скелету присоединен азотсодержащий гетероцикл (рис. 10). Такие соединения называют гликоалкалоидами. К ним относится хорошо известный соланин, накапливающийся в ягодах и озелененных после выставления на свет клубнях картофеля. В зараженных растениях от растворимого сапонина с помощью ферментов гликози-даз отщепляются сахара и терпеноидный агликон проявляет свою высокую токсичность.
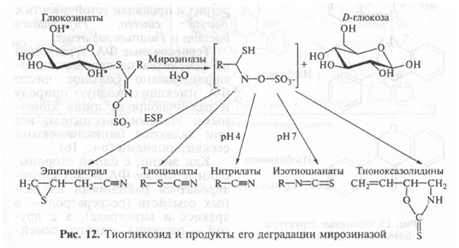
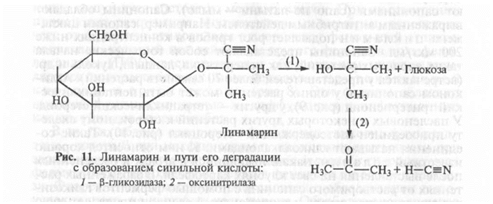
Цианогенные гликозиды. У ряда растений (проса, лядвенца, сорго и др.) содержатся своеобразные гликозиды (дурринин, линама-рин и др.), у которых при повреждении мембран сначала (3-глико-зидаза отщепляет глюкозу, а затем под действием другого фермента — оксинитрилазы — образуется сильнейший дыхательный яд — синильная кислота (рис. 11).
Тиогликозиды. У этих соединений агликон соединен с сахаром не через кислород, а через серу. В результате воздействия специфических гликозидаз (миронозидазы и др.) образуется ряд летучих высокотоксичных соединений (рис. 12). Тиогликозиды, или гл и -ценоляты, присущи растениям из семейства крестоцветных, которым они придают специфический запах от слабого у капусты до очень резкого у редьки и хрена, они также обнаружены у растений некоторых других семейств.
Фитоалексины. В 1940 г. немецкие фитопатологи Мюллер и Бергер показали, что в клубнях устойчивых к фитофторозу сортов картофеля после заражения накапливаются токсические вещества, которые убивают мицелий патогена, развивающийся в тканях растения. Эти вещества были названы фитоалексинами (ФА). Согласно предложенной авторами концепции, ФА —низкомолекулярные растительные антибиотики, которые отсутствуют
 |
 |
и в интактных, и в механически поврежденных тканях, но синтезируются в ответ на заражение невирулентными видами и штаммами патогенных грибов. Они локализованы в зоне заражения и не мигрируют в незараженные ткани.
 |
Методы изучения ФА. Заражение срезов клубней картофеля, как это было сделано в первых опытах Мюллера, оказалось не вполне корректным методом изучения ФА, так как при разрезании клубня происходит повреждение клеток и образование описанных ранее стрессовых метаболитов — антиципинов. (Поэтому в лаборатории Л. В. Метлицкого на поверхности среза клубня специальной ложечкой вырезают отверстия, промывают их водопроводной водой для удаления стрессовых метаболитов, подсушивают и затем заполняют каплями со спорами.) Нанесение капель со спорами на поверхность листьев также неинформативно, поскольку кутикула препятствует обмену метаболитов в системе паразит-хозяин и накоплению ФА. Поэтому в качестве моделей для изучения ФА Мюллер выбрал плоды гороха и фасоли. Он извлекал из створок бобов семена и в углубления во внутренней части плодов (семенные камеры) наносил водную суспензию спор гриба Monilia fructicola (возбудитель плодовой гнили яблони), непатогенного для бобовых. Таким образом, использовалась неповрежденная ткань, лишенная кутикулярного слоя. В опытах было два контроля: в семенные камеры вносили капли воды без спор (контроль на наличие неспецифических антиципинов); суспензию спор наносили на предметные стекла во влажной камере (контроль на наличие автоингибиторов, выделяемых прорастающими спорами гриба). После определенной экспозиции капли собирали, отделяли от спор центрифугированием и в полученной жидкости проращивали свежую порцию спор М. fructicola. Ингибирование прорастания спор в опыте по сравнению с контролями свидетельствовало о диффузии ФА в инфекционную каплю. Кроме створок плодов бобовых растений использовали плоды и других растений, например перца, которые разрезали вдоль, извлекали семена и заливали суспензиями спор разных грибов. Для изучения ФА, образующихся в ответ на системную инфекцию грибами, локализованными в ксилеме, зараженные растения обрезали на пенек и собирали пасоку (жидкость, поднимаемую вследствие корневого давления). Так были изучены ФА хлопчатника, зараженного вертициллезным вилтом.
. Химическая природа ФА. В настоящее время описана структура нескольких сотен ФА из различных однодольных и двудольных растений. По химической природе их разделяют на несколько групп.
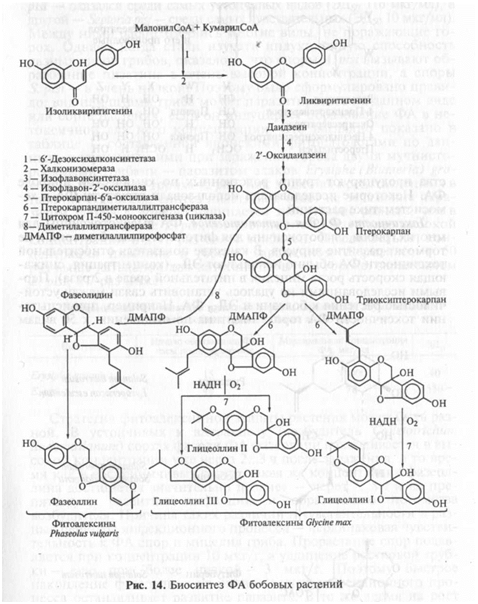
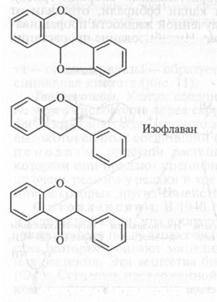
Фенольные ФА. Большинство бобовых растений образует сложные фенольные ФА — изофлаваны (киевитон фасоли), изофлава-ноны (веститол лядвенца, сативан люцерны), птерокарпаны (пизатин гороха, фазеоллин фасоли, глицеоллин сои, медикарпин люцерны). Общая структура этих групп соединений показана на рисунке 13. Как видно, они содержат два фенольных кольца, синтезируемых разными метаболическими путями (через шикимовую и через уксусную кислоты). Активность фермента, соединяющего эти кольца — халконсинтетазы, увеличивается вслед за заражением. Биосинтез ФА бобовых показан на рисунке 14.
У арахиса и винограда два фенольных кольца ФА ресвератро-ла (рис. 15) соединяет другой фермент— стильбенсинтетаза (СТС). Гены, контролирующие синтез СТС, были методами генной инженерии перенесены из винограда Птерокарпан в табак, томаты и картофель, которые стали продуцировать ресве-ратрол и проявили устойчивость к Botrytis cinerea, Phytophthora infestans и Fusarium sulfureum.
Терпеноидные ФА. У растений из семейства пасленовых идентифицировано большое число ФА, имеющих сходную природу и различающихся лишь конечными этапами биосинтеза; все они являются бициклическими сесквитерпенами (рис. 16). Изофлаванон Как видно, с одной стороны, один и тот же ФА может синтезироваться растениями из разных семейств (ресвератрол — в арахисе и винограде), а с другой – растения одного семейства продуцируют группу родственных по химической природе ФА. Некоторые исследователи использовали структуру ФА в хемосистематике растений.
Рис. 13. Основные структуры флавоновых фитоалексинов
|
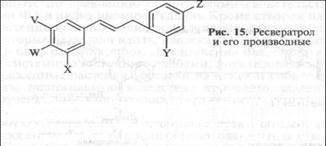
Ресвератрол ОН Н ОН Н ОН
4-Пренилресвератрол ОН Прения ОН Н ОН
Оксиресвератрол ОН Н ОН ОН ОН
4-Пренилоксиресвератрол ОН Прения ОН ОН ОН
Птеростильбен ОСН3 Н ОСН3 Н ОН
Токсичность ФА для фитопатогенов. ФА высокотоксичны для многих грибов, слаботоксичны для фитопатогенных бактерий и не тормозят развитие вирусов. В качестве показателя относительной токсичности ФА обычно используют ЭД50 (концентрация, снижающая скорость роста мицелия в питательной среде в 2 раза). Первыми исследованиями не удалось установить связи между устойчивостью растения к болезни и ЭД50 ФА. Например, при испытании токсичности ФА гороха пизатина по отношению к 50 видам



грибов было показано, что один из паразитов гороха — Ascochyta pisi — оказался среди самых устойчивых видов (ЭД50 116 мкг/мл), а другой — Septoriapisi— среди самых чувствительных (ЭД50 10 мкг/мл). Между ними расположились многие виды, не поражающие горох. Однако когда стали изучать* индукционную способность разных видов грибов, оказалось, что споры A. pisi вызывают образование пизатина в очень высокой концентрации, а споры S. pisi — в очень низкой. Поэтому было сформулировано правило: вид или штамм гриба может паразитировать на данном виде или сорте растения, если он индуцирует образование ФА в нетоксичной для него концентрации. Это наглядно показано в таблице 1, составленной японскими фитопатологами по данным, полученным ими при заражении гороха двумя мучнисто-росяными грибами — паразитом злаков Erysiphe (Blumeria) gra-minis и паразитом гороха Е. pisi. Как видно из представленных в таблице 1 данных, синтез пизатина в ответ на заражение паразитом злаков Е. graminis начинается значительно раньше, чем в ответ на заражение Е. pisi, он достигает в 25 раз более высокой концентрации и в 13 раз более токсичен. Следовательно, концентрация пизатина в горохе, зараженном Е. graminis, в 50 раз выше летальной для патогена дозы, а в горохе, зараженном Е. pisi, в 7 раз ниже.








