 2013-12-31
2013-12-31 1144
1144Устойчивость растений обусловлена комплексным действием разных факторов. Механизмы или факторы устойчивости можно подразделить на две группы:
* факторы, действующие до заражения (прединфекционные);
* факторы, действующие после заражения (постинфекционные).
Факторы (механизмы) устойчивости первой группы присутствуют в растении независимо от поражения, второй - индуцируются
возбудителями (к этой группе относится и изменение активности генов). Среди факторов, действующих до заражения, можно выделить анатомо-морфологические, физические и химические.
Габитус растений. Начальный этап заболевания (прорастание спор) в большинстве случаев требует наличия капельно-жидкой влаги на поверхности растений. Прорастанию спор благоприятствует также высокая влажность воздуха. В связи с этим габитус растения может создавать благоприятные условия для заражения, если он способствует сохранению высокой влажности окружающего воздуха и позволяет каплям воды задерживаться на поверхности надземных органов растения. Так, сорта картофеля с рыхлым кустом менее подвержены заражению фитофторозом, чем сорта с плотным кустом, хотя при искусственном заражении листьев связи между габитусом куста и поражаемостью нет. Отсутствие различий объясняется тем, что период увлажнения, необходимый для заражения надземных частей растений, у сортов с раскидистым кустом короче, чем у сортов с плотным.
Опушенность листьев. Сорта картофеля с сильноопушенными листьями менее подвержены вирусным болезням, чем сорта со слабым опушением. Это обусловлено более слабым посещением опушенных растений тлями — переносчиками вирусов из-за менее благоприятных условий для питания.
Толстый кутикулярный слой. Фактором, препятствующим поражению растения различными заболеваниями, может служить толщина кутикулярного слоя (рис. 1). Так, у сортов крыжовника, устойчивых к мучнистой росе, толщина кутикулы молодых листьев 1,05...1,26 мкм, а у восприимчивых сортов — 0,51...0,64 мкм. У сортов барбариса, неустойчивых к ржавчине, толщина кутикулярного слоя у листьев составляет вместе с эпидермисом 0,82 мкм, тогда как у устойчивых сортов — 1,75 мкм. Субстанции, способствующие отвердению эпидермальных слоев растений, например лигнин, кремниевая кислота, кальций и др., повышают сопротивляемость ткани и препятствуют проникновению грибов. По сравнению с
надземными эпидермальными тканями ризодермис, как правило, является более легким барьером для проникновения многих видов грибов в корень.
Строение и расположение устьиц и чечевичек. Возможность заражения растений некоторыми патогенами зависит от числа и строения устьиц и чечевичек. Закрытые устьица и чечевички задерживают заражение растений патогенами.
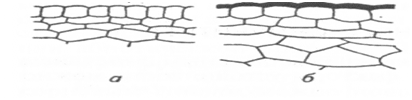
Рис. 1. Разрез через наружные слои.
восприимчивого (а) и устойчивого
(б) и к грибу Botrytis cinerea растений
 |
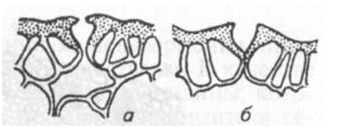
Виды и сорта растении с более редким расположением устьиц на листьях менее поражаются грибной и бактериальной инфекцией. Мандарин более устойчив к возбудителю бактериального рака Xanthomonas citri, чем грейпфрут, потому что наружные стенки устьиц мандарина снабжены выступами, препятствующими проникновению в подустичную щель капель жидкости с находящимися в них клетками бактерий. У восприимчивых сортов грейпфрута таких выступов нет (рис. 2).
Восковой налет. Наличие его придает поверхности органов растения гидрофобные свойства, что затрудняет ее смачивание и препятствует прорастанию спор. Восковой налет задерживает поступление питательных веществ, которые могут способствовать росту возбудителей на поверхности растения. Каплеудерживающую способность листьев пшеницы часто используют как один из показателей ее устойчивости к бурой листовой ржавчине.
Особенности строения цветка. Длина пыльника, характер цветения (открытое или закрытое), его продолжительность и т. д. играют роль в устойчивости к патогенам, заражающим растения во время цветения (например, возбудителям пыльной головни пшеницы, ржи, ячменя и др.).
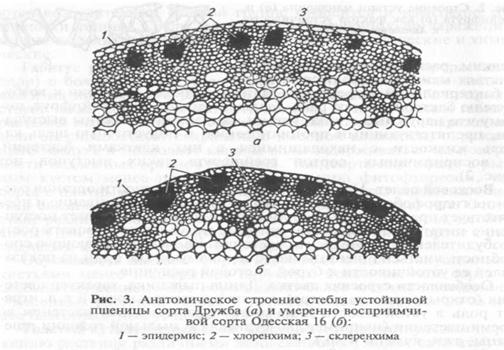
Анатомические особенности внутренних тканей оказывают существенное влияние на устойчивость растений. Например, восприимчивые к стеблевой ржавчине сорта пшеницы имеют развитую хлоренхиму, которая залегает под эпидермисом широкими тяжами, в то время как у устойчивых сортов эта ткань имеет вид островков, недоступных для колонизации патогеном (рис. 3). Одревесневшая эндодерма корней, особенно у однодольных, представляет для многих видов грибов, например рода Fusarium, физический барьер, препятствующий их проникновению в растение.
Пробковый слой. Играет важную роль в устойчивости растений на этапе внедрения патогенов. Например, грибы родов Fusarium, Phytophthora infestans и некоторые бактерии слабее поражают клубни картофеля с хорошо развитым пробковым слоем.
Химический состав растений. Относится к важным факторам пассивного иммунитета. Устойчивость иногда обусловливается отсутствием или низкой концентрацией в тканях растений необходимых для патогена питательных веществ. Часто некоторые химические соединения, содержащиеся в растении, подавляют развитие патогена.
Устойчивость к патогенам может быть обусловлена как отсутствием в растении веществ, необходимых для их развития, так и
содержанием токсичных для него веществ. Вакуолизация снижает содержание протоплазмы в клетках, а следовательно, ухудшает условия для питания облигатно-биотрофных грибов.
Действие всевозможных специфических патотоксинов коррелирует с присутствием природных рецепторов на клетках растения-хозяина. Устойчивость к таким патотоксинам часто объясняется отсутствием этих рецепторов у растения-хозяина. Содержание в растениях антибиотических веществ, таких, как фенолы, флавоны, сапонины, лактоны, танины и др., связывают с устойчивостью к возбудителям грибных заболеваний. Так, сапонины оказывают токсичное действие на фитопатогенные грибы, содержащие в своих мембранах стерины. В случае заражения растения овса возбудителем офиоболеза пшеницы (Ophiobolus graminis var. tritici) содержащийся в его корнях сапонин авенацин активируется и гриб погибает. Если же возбудитель офиоболеза овса (Op. graminis var. avena), содержащий ферменты, которые могут инактивировать сапонины, поражает растения овса, то инфекция сохраняется, заражение и развитие болезни становится возможным.
Часто патогенность возбудителей болезней снижается вследствие того, что вырабатываемые патогенами и используемые ими для проникновения и заселения растений-хозяев ферменты инак-тивируются содержащимися в растениях веществами. В качестве примера таких веществ можно назвать проантоцианидины, которые инактивируют протеолитические ферменты возбудителя серой гнили (Botrytis cinerea) в незрелых плодах земляники, и ингибиторы протеаз, которые широко распространены в растительных тканях.
Углеводы. Одни патогены, такие, как возбудители ржавчины и мучнистой росы, лучше развиваются при высоких концентрациях Сахаров, а другие, например возбудитель бурой альтернариозной пятнистости томата, — при низких. Очевидно, что растение становится устойчивым к патогенам, требующим высоких концентраций Сахаров, в том случае, когда их содержание в нем падает до определенного уровня.
Факультативные паразиты легче поражают ткани, обедненные углеводами (что может служить одним из признаков ослабления растений). Облигатные паразиты развиваются только на живых тканях, сохраняющих фотосинтетическую активность. Вероятно, поэтому восприимчивые к ржавчине растения пшеницы не заражаются паразитом после выдерживания их в темноте.
Белки и продукты их распада. Устойчивость растительных тканей к факультативным паразитам положительно коррелирует с высоким содержанием белка и относительно низким содержанием более подвижных соединений азота. Например, в тканях устойчивого сорта капусты Амагер содержится значительно больше белка, в то время как восприимчивому сорту Номер первый свойственно высокое содержание аминокислот (Арциховская, 1956). Однако недостаток растворимых азотистых соединений вряд ли может оказывать существенное влияние на поражаемость, поскольку факультативные паразиты способны синтезировать активные ферменты, расщепляющие белковые соединения растительных тканей до усвояемых паразитами продуктов.
Обмен веществ облигатных паразитов оптимально приспособлен к обмену веществ растения-хозяина. Основной фактор химического состава устойчивости растений к облигатным паразитам — несоответствие растения-хозяина пищевым потребностям паразита. Восприимчивость растений к возбудителям мучнистой росы и ржавчины обусловлена наличием в них специфических веществ, необходимых для питания паразита.
Органические кислоты. Большинство патогенов, главным образом грибов, выдерживают высокие концентрации кислот и способны развиваться в широких пределах рН. Более существенную роль играет кислотность тканей в случае взаимоотношения растений с возбудителями бактериальных заболеваний, поскольку бактерии, в том числе и фитопатогенные, весьма чувствительны к понижению рН среды. Так, возбудитель бактериальной пятнистости
{Xanthomonas vesicatoria) легко заражает зеленые плоды томата, рН клеточного сока у которых выше 5, в то время как спелые плоды с рН 4...5 полностью устойчивы к заболеванию.
Органические кислоты могут воздействовать на паразита не непосредственно, а путем усиления действия токсинов, содержащихся в тканях растений. Так, экстракты из крестоцветных содержат фенольные вещества (они обладают слабыми кислотными свойствами), токсичные для Colletotrichum circinans. Действие этих веществ зависит от концентрации водородных ионов. Максимальная токсичность гидрохинона достигается при рН 4, а катехола — при рН 5.
Многие фитопатогены могут развиваться лишь при наличии в питательной среде физиологически активных веществ типа витаминов, так, бедные тиамином сорта зерновых культур иммунны к возбудителю твердой головни Tilletia tritici.
Некоторые красящие вещества, например антоцианы, по своей химической структуре относятся к гликозидам. Установлено, что многие окрашенные сорта некоторых культур поражаются слабее, чем бесцветные.
Фитонциды (фитоантицепины). По определению Б. П. Токина, фитонциды — летучие вещества растений, ингибирующие рост и развитие микроорганизмов. Фитонцидной активностью обладают различные химические соединения, главным образом низкомолекулярные (органические кислоты, альдегиды, монотерпены, простые фенолы).
По объекту действия Токин разделил фитонциды на три группы:
• бактерицидные, фунгицидные и протистоцидные;
• стимулирующие или тормозящие прорастание пыльцы, рост и развитие растений;
• токсичные для насекомых, клещей, червей и других организмов.
Показано, что фитонцидное действие растений на специализированных к ним паразитов, как правило, выражено слабо. В большинстве случаев оно распространяется на микроорганизмы, не поражающие данное растение. Наиболее мощным фитонцидным действием на возбудителей ряда бактериальных болезней, например томата, обладают чеснок, лук и мята. Фитонциды присущи всем видам растений, однако их активность неодинакова. Она меняется в зависимости от вида, сорта и возраста растений, а также от времени дня, фазы развития растений, погодных условий и т. п. При механическом повреждении тканей количество выделяемых фитонцидов резко возрастает. Фитонциды более эффективно действуют на сапротрофных фи-топатогенов, чем на облигатных паразитов. Фитонциды могут служить одним. из факторов врожденного неспецифического иммунитета растений.
2.2. ФАКТОРЫ АКТИВНОГО ИММУНИТЕТА
Активный иммунитет растений проявляется в виде комплекса ответных реакций на воздействие патогенных организмов или продуктов их жизнедеятельности. Сущность активного иммунитета сводится к определенным биохимическим процессам. Как отмечал И. И. Мечников, существует два типа защитных реакций организма на инфицирование. В одних случаях они направлены на обезвреживание токсических выделений патогена, в других, и главным образом, — непосредственно на подавление самого возбудителя. Учение Мечникова о двух типах защитных реакций в полной мере подтверждают данные, полученные при изучении иммунитета растений. Характер защитных реакций растений определяется в основном типом паразитизма и способом питания патогена.
К факторам активного иммунитета относят реакцию сверхчувствительности, активацию и перестройку деятельности ферментных систем, образование фитоалексинов, фагоцитоз.
Результаты активных защитных реакций могут проявляться либо в локализации патогена, либо в замедлении его распространения в растительных тканях, либо в его гибели и предотвращении болезни растения.
Обычно вопрос о совместимости или несовместимости возбудителя и растения-хозяина решается только после проникновения возбудителя в растение. Как правило, возбудитель грибной или вирусной природы проникает в растение независимо от того, является оно хозяином или нет. Лишь после этого на уровне плазмалеммы клетки происходят генетически регулируемые процессы узнавания. При этом экзогенные сигналы, или элиситоры, связываются пока мало исследованными рецепторами. Элиситоры даже в очень малых концентрациях способны вызывать каскад защитных реакций растения-хозяина еще на стадии заражения растения возбудителем. Действие их неспецифично, они индуцируют общую активацию устойчивости. Установлена структура лишь некоторых из них, например олигомеров хитина, хитоназы и глюканов. Это арахидоновая и эйкозопентаэновая кислоты, некоторые гликопротеиды из грибов, олигогалактурониды, системин, салициловая, изоникотиновая, линоленовая и жасмоновая кислоты у растений.
Экзогенные элиситоры преобразуются на плазмалемме в эндогенные сигналы. Первой реакцией при этом, вероятно, является фосфорилирование протеинов, в котором участвуют разные ферменты (фосфолипаза, протеинкиназа, плазмалемма—АТФаза). Эти эндогенные сигнальные молекулы вызывают у растений все дальнейшие защитные реакции. Через цитоплазму эти молекулы попадают в ядро клетки и активируют гены устойчивости. Сначала происходит индукция или стимуляция транскрипции и трансляции РНК, что приводит к образованию ферментов. Они катализируют синтез генных продуктов, связанных с защитными реакциями. Сигналы могут действовать только в отдельных или соседних клетках либо путем транслокации попадать в более далекие свободные от поражения части растения и индуцировать там защитные реакции. Последовательность этих реакций называют сигнальными цепями, под которыми понимают совокупность всех реакций от рецепции сигнала до фенотипического проявления реакции. Последовательность таких реакций пока не установлена, поэтому сигнальные цепи носят частично гипотетический характер.
Результатом активизации устойчивости может быть образование структур (раневой перидермы, лигнина), с помощью которых растение пытается предотвратить заселение и инкапсулировать возбудителя в местах его проникновения. При развитии реакции несовместимости у двудольных растений наблюдается усиленное накопление в клеточных оболочках гликопротеидов. У однодольных (ячменя) при поражении мучнистой росой в клеточной оболочке накапливается белок тионин.
Сверхчувствительность. Один из наиболее эффективных и распространенных механизмов устойчивости растений — реакция сверхчувствительности. Она индуцируется многими возбудителями и протекает очень быстро: происходит гибель клеток хозяина в местах проникновения патогена. Мертвые клетки становятся для патогена барьером, изолирующим его от живой ткани (рис. 4).
Реакция сверхчувствительности возникает в ответ на заражение устойчивых сортов биотрофами, поскольку эта группа паразитов способна использовать для питания только живые клетки. Отмирание окружающих тканей ведет к голоданию биотрофов, а затем и к их гибели. Факультативные паразиты (некротрофы) способны продолжать развитие в мертвых тканях как сапротрофы. Однако в ряде случаев реакция сверхчувствительности является важным фактором иммунитета и к факультативным паразитам, гибель которых наступает в результате отравления образующимися токсинами. В данном случае защитная реакция растения направлена непосредственно против патогена.
Внешне реакция сверхчувствительности проявляется в виде некрозов, которые наиболее четко выражены при внедрении в растение облигатных паразитов и факультативных сапротрофов, оптимально приспособленных к обмену веществ растения-хозяина.
Установлено, что первые этапы патологического процесса (проникновение возбудителя в ткани растения-хозяина) у устойчивых и восприимчивых сортов протекают одинаково. На последующих этапах патологического процесса реакция сортов резко меняется. У восприимчивых сортов гриб внедряется в клетки и питается за счет растения. Клетки, в которые проник гриб, продолжают функционировать. При этом наблюдается как бы симбиоз патогенного организма с высшим растением. У устойчивых сортов после проникновения гифы в клетку ее ядро и все содержимое начинают темнеть и погибают вместе с гифой.
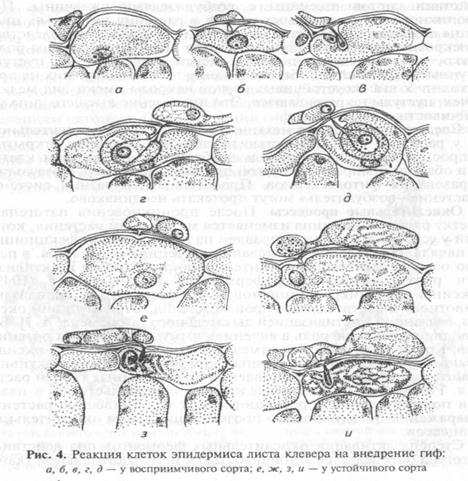
В некоторых случаях реакция сверхчувствительности не ограничивается гибелью только инфицированных клеток: отмирают и граничащие с ними клетки. Гибель клетки вместе с паразитом прерывает инфекционный процесс и предотвращает дальнейшее развитие болезни. Скорость и интенсивность реакции сверхчувствительности при разных сочетаниях хозяина и патогена неодинакова: чем устойчивее сорт, тем быстрее развивается реакция сверхчувствительности. Так, красный клевер, устойчивый к Erysiphe hiligoni, мгновенно реагирует на вторжение паразита. При этом происходит быстрая дезорганизация и гибель пораженных и некоторых примыкающих к ним клеток, а также гиф гриба. В случае восприимчивых разновидностей такой реакции не наблюдалось. Аналогичные явления были обнаружены при взаимодействии сортов пшеницы с возбудителями ржавчины. При проникновении ржавчинных грибов в растения на листьях пшеницы появляются хлоротичные пятна, переходящие впоследствии в некротические. У устойчивых сортов некрозы выражены более четко, чем у восприимчивых. У неустойчивых сортов пустулы крупные, бархатистее, обесцвечивание тканей вокруг них не происходит. У высокоустойчивых сортов некрозы имеют вид мелких точек, пустулы не развиваются. Это проявление высшего типа устойчивости — иммунитета.
Следует отметить, что механизм реакции сверхчувствительности у растений пока полностью не выяснен. Остается открытым вопрос, обусловлена ли гибель возбудителя отмиранием клетки или оба они отмирают по другой причине, например, в результате образования фитоалексинов. Процессы эти в разных системах «растение—возбудитель» могут протекать неодинаково.
Окислительные процессы. После проникновения патогена в клетку растения-хозяина изменяется метаболизм растения, который у устойчивых форм направлен на подавление инфекционного начала. Проявление заболевания растения заключается, в первую очередь, в повышении интенсивности дыхания и активизации ряда окислительных ферментов. По А. Н. Баху (1949), токсины ряда микроорганизмов могут разрушаться оксидазами животного и растительного происхождения. Этой функции оксидаз, связанной с активизацией дыхательного газообмена, А. Н. Бах отводил решающую роль в явлениях иммунитета живых организмов. Кроме того, А. Н. Бах отмечал специфическую роль оксидаз в защитных реакциях, эта группа ферментов принимает активное участие в процессах восстановления поврежденных тканей растения. Результаты многих исследований свидетельствуют о наличии положительной корреляции между устойчивостью растения к заражению и активностью протекающих в нем окислительных процессов.
Степень активации окислительных ферментов под действием патогена определяется устойчивостью к нему растительной ткани.
У устойчивых сортов активная реакция на заражение сопровождается усилением энергетического обмена (рис. 5). У восприимчивых растений под влиянием паразита наблюдаются распад окислительных систем и нарушение отдельных звеньев обмена веществ (рис. 6). Под влиянием окислительной системы растения происходит снижение активности гидролитических ферментов патогена, нейтрализация его токсинов. Защитные реакции, направленные на подавление ферментов паразита, называются антиферментными, а направленные на разрушение токсинов паразита — антитоксическими.
 |
На инактивацию ферментов паразита или на ингибирование их синтеза могут оказывать влияние фенольные соединения, аминокислоты или катионы металлов, от которых зависит активность ряда ферментов. Фенолы и продукты их окисления подавляют активность многих ферментов. Устойчивость может быть также связана с отсутствием в клетках растения субстрата для ферментов, без которых невозможно заселение хозяина возбудителем. Катионы кальция, бария и магния могут влиять на активность пектолитических, а возможно, и других ферментов и благодаря этому играть определенную роль в устойчивости растений. Устойчивость к инфекции определяется устойчивостью клеток к токсическим выделениям патогена. При внедрении гиф Botrytis cinerea в ткани неустойчивых сортов капусты наблюдается быстро распространяющееся потемнение тканей. Мицелий гриба не пораженной ткани можно обнаружить уже в первые дни заражения. У устойчивых сортов потемнение тканей
 |
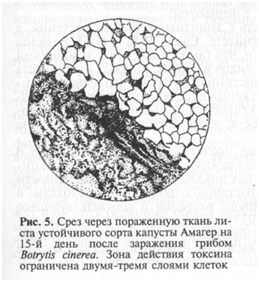
и налет конидиеносцев возбудителей заболевания появляются лишь на восьмые сутки. Различия в площади, занятой мицелием, и площади, потемневшей от действия токсина, у восприимчивого сорта и почти полное совпадение этих показателей у устойчивого сорта свидетельствуют о том, что именно инактивация токсинов определяет защиту от патогена.
Деятельность окислительных ферментов как факторов активного иммунитета растений может проявляться и в процессах, ведущих к образованию между паразитом и растительной тканью защитного слоя клеток, который изолирует патоген. При заражении клубней картофеля возбудителем обыкновенной парши (Streptomyces scabies) у устойчивого сорта вокруг внедрившегося паразита быстро возникает слой пробки, тогда как у восприимчивого сорта развитие пробки происходит медленнее и оказывается неэффективным. Успешное образование барьера из пробки приводит к локализации очагов болезни. Барьер из пробки может препятствовать распространению токсинов из пораженных тканей. Таким образом, окислительные ферменты являются активными участниками биохимических процессов, происходящих в результате взаимодействия растений и паразита. Интенсивность и характер этих процессов, их влияние на конечный исход заражения зависят от присущих хозяину и патогену особенностей обмена веществ, а также от тех изменений, которые возникают в обмене веществ в результате их взаимодействия. Эти сдвиги в обмене обусловлены изменениями в деятельности ферментов.
Белковый обмен. Все активные защитные реакции растения связаны с существенными изменениями в обмене веществ как растения, так и паразита. Регуляторами этих изменений являются нуклеиновые кислоты, белки и ферменты. Белки играют ведущую роль в защитных реакциях растения, а следовательно, и в фитоиммунитете. Еще в 1940 г. Н. И. Вавилов подчеркивал возможность того, что по мере изучения химической природы белков, различной у видов и сортов, природа иммунитета станет более понятной. Он констатировал также, что наряду с усилением дыхания процесс заражения нередко сопровождается возрастанием ферментативной активности и появлением новых ферментов, белков и других веществ, ранее отсутствующих у хозяина и паразита.
Результаты многочисленных исследований свидетельствуют о том, что белковый обмен играет важную роль в защитных реакциях растений. Одним из основных показателей приспособленности патогенов к обмену веществ растения-хозяина служит сходство в строении белков восприимчивого растения и соответствующего паразита. Т. И. Федотова в 1935 г. обнаружила сходство иммунологических свойств белков возбудителей болезней и их растений-хозяев, а в 1966 г. ей удалось выделить иммунологически сходные компоненты белков, связанные с проявлением паразитизма у возбудителей. Предположения о существовании общих антигенов у паразита и восприимчивого растения послужили основой для работ по изучению роли белков в иммунитете растений. М. С. Дунин с сотрудниками изучали устойчивость растений к заболеваниям с помощью серологического метода, Основанного на учете антигенной специфичности белков устойчивых и восприимчивых к заболеванию форм.
Большой интерес к проблеме совместимости растения-хозяина и патогена проявлял Стаман (1968). Он предположил, что в случае поражаемое™ растения патогеном в них синтезируются ферменты со сходными свойствами. При взаимодействии восприимчивого растения и патогена происходит обмен между ферментными системами и образование своего рода гибридных ферментов, отличающихся по свойствам от ферментов растения и паразита. От характера изменений в обмене веществ растения-хозяина зависит исход взаимодействия, т. е. гибель паразита или поражение растения. Стаман считал, что успешное развитие болезни возможно только в случае сходства ферментных систем растения-хозяина и патогена.
О наличии связи между особенностями белкового обмена растения-хозяина и его восприимчивостью или устойчивостью к тому или иному заболеванию свидетельствуют результаты многочисленных работ отечественных и зарубежных исследователей. Так, Д. В. Липсиц (1964) установил, что белки устойчивых и восприимчивых к раку сортов картофеля различаются по некоторым физико-химическим показателям, например аминокислотному составу, иммунохимическим свойствам и т.д. Молекулярная структура белков неустойчивого сорта менее прочна, чем у устойчивого, поэтому при внедрении патогена они подвергаются изменениям, что приводит к повышению ферментативной активности и вызывает активную ростовую реакцию. В результате усиленного деления клеток происходит образование наростов. В тканях устойчивых сортов подобной реакции не наблюдается. К тому же белки устойчивых сортов более устойчивы к действию протео-литических ферментов, чем белки восприимчивых. Реакция растений на заражение может проявляться также -в новообразовании белков с самыми разнообразными функциями, поскольку они не только участвуют во всех звеньях метаболизма, но и определяют фитотипические свойства каждого организма, особенности его взаимодействия с паразитом. Изменения свойств белков, происходящие в растениях при заражении, влияют на ход всех обменных процессов клеток, а следовательно, и на результат заражения.
Были также высказаны предположения о том, что хозяин может изменить белковый обмен паразита, и о возможном влиянии аминокислот на устойчивость растений к патогенам. Очевидно, аминокислоты, изменяя метаболизм растения-хозяина, способствуют появлению фунгицидных или фунгистатических веществ либо вызывают какие-то изменения, в результате которых ткани растения становятся менее пригодными для питания патогена (Андель, 1966). Следовательно, ни одна из рассмотренных защитных реакций (активация дыхания и других окислительных процессов) не может осуществляться без участия белков и их мономеров, без предварительного воздействия на них.
Фитоалексины. Это низкомолекулярные антибиотические вещества, синтезирующиеся в растении в результате взаимодействия продуктов метаболизма растения-хозяина и возбудителя. Фитоалексины ингибируют развитие микроорганизмов, попадающих на растение. Впервые образование растениями фитоалексинов в ответ на инфекцию обнаружил К. О. Мюллер (1939), который изучал образование некрозов при заражении клубней картофеля грибом Phytophthora infestans. В нашей стране теория фитоалексинов получила дальнейшее развитие в многочисленных исследованиях Л. В. Метлицкого, О. Л. Озерецковской и др.
Установлено, что синтез фитоалексинов живыми клетками — реакция не только на их поражение возбудителями болезней, но и на повреждение, воздействие других раздражителей, стресс и т. д. (рис. 7). Это временный процесс, который зависит от внешних условий (температуры, освещения, питания и т. д.). К настоящему времени из растений разных ботанических семейств выделено и идентифицировано более 200 фитоалексинов, из бобовых, пасленовых и др. — более 20, меньше всего их обнаружено у злаковых. При взаимодействии патогена с растением может синтезироваться несколько фитоалексинов. Так, при заражении Ph. infestans в клубнях картофеля образуются три фитоалексина (ришитин, люби-мин и фитуберин), а после инфицирования бобов вирусом некроза табака — четыре. Способность растения продуцировать не один, а несколько фитоалексинов позволяет ему успешно противостоять разным патогенам.
 |
По химической природе фитоалексины относятся к разным классам соединений вторичного обмена, таким, как полиацетилены (вайероновая кислота), полифенблы (госсипол), изофлавонои-ды (пизатин, фазеоллин, глицеоллин), терпеноиды (ришитин, капсдиол) и стилбены (ресвератрол). Фитоалексины характерны для вида, рода высшего растения. Фитоалексины растений близких таксонов сходны по своей химической структуре. Наиболее известные фитоалексины фазеоллин (С2оН1804) из фасоли (его индукторы Botrytis cinerea, Monilia fructigena, Uromyces phaseoli, Colletotrichum spp.), пизатин (С17Н140б) из гороха (его индукторы Fusarium solani f. sp. pisi, F. ssp. phaseoli), ришитин (С14Н2202) из томата, клубней картофеля (их индукторы Fulvia fulva, Ph. infestans) и Др.
Фитоалексины, по-видимому, неспецифичны по отношению к патогену, так как синтез одного и того же фитоалексина, характерного для данного растения, может быть вызван разными патогенами. Установлено, что грибы, паразитирующие на растении, более устойчивы к фитоалексинам, вырабатываемым данным растением, чем непатогенные виды. Так, гриб Ascochyta pisi, паразитирующий на горохе, менее чувствителен к пизатину и более чувствителен к фазеоллину из фасоли, на которой он не паразитирует.
Доказано, что большинство фитоалексинов обладает фунгиста-тическим действием, хотя количественных данных по этому показателю пока недостаточно. Так, зерновые культуры (пшеница, кукуруза, рожь) содержат глюкозиды дегидрооксибензоксазолиноны и фермент глюкозидазу, взаимодействие которых в пораженных тканях приводит к образованию аглюконов — веществ с фунги-цидными свойствами.
В некоторых случаях фитоалексины, несомненно, играют существенную роль в защитных реакциях. Однако не всегда и не во всех растениях, в которых образуются, они служат препятствием для фитопатогенных грибов. Например, доказано участие фитоалексина глицеоллина в устойчивости сои к грибу Phytophthora megosperma var. sojae, однако в старых растениях, когда снижается его синтез, устойчивость растений повышается. При повышенной температуре восприимчивые сорта сои синтезируют большое количество глицеоллина, однако не приобретают при этом устойчивости. Вероятно, фитоалексины, как и другие токсичные вещества, — лишь один из компонентов в чрезвычайно сложной системе реакций, обусловливающих устойчивость растений. Более подробно материалы, касающиеся раздела «Факторы активного иммунитета», будут рассмотрены в главе «Генетика, биохимия и молекулярная биология иммунитета растений».
Фагоцитоз. Этот процесс был впервые обнаружен у животных И. И. Мечниковым. Он же создал фагоцитарную теорию защиты
животного организма от инфекции. Согласно этой теории, у животных есть специальные клетки (фагоциты), обладающие способностью захватывать паразита и переваривать его с помощью ферментов. У растений в отличие от животных нет фагоцитов. Однако еще в начале XX в. появились сообщения о том, что у растений обнаружен сходный процесс внутриклеточного переваривания, получивший название фагоцитоза. Поскольку такая способность у растений проявляется лишь в отношении патогенных организмов, она была отнесена,< факторам активного иммунитета. Фагоцитоз четко проявляется в случае эндотрофной микоризы, когда гриб развивается внутри корней растения и лишь небольшая его часть проникает из корней в почву. Эндотрофная микориза развивается следующим образом: гифы гриба проникают через корневые волоски в эпидермальные клетки, а затем в клетки первичной коры, в которых происходят защитные реакции, вызывающие замедление распространения в них мицелия: гифы либо становятся клубочкообразными, либо у них разветвляются верхушки с образованием так называемых арбускул. Внутриклеточные клубочки гиф постепенно перевариваются клетками и исчезают. Арбускулы теряют способность к росту, и дальнейшее распространение мицелия по клеткам первичной коры прекращается. Такие взаимоотношения обнаружены на микоризах деревьев, злаков и других растений. У злаков, например, Fusarium oxysporum образует эндотрофную микоризу. При этом иногда наблюдается частичное или полное переваривание гиф в клетках корня. Очевидно, благодаря фагоцитарным свойствам клетки сдерживают распространение гриба, не дают ему перейти к паразитическому образу жизни, однако не уничтожают его полностью. Подобное равновесие между корнями злаков и грибом непостоянно и зависит от факторов внешней среды.

