2013-12-31
2013-12-31 858
858N= 2",
где п — число растений-дифференциаторов.
Основание степени равно двум, потому что имеются только две возможные реакции: совместимости и сверхчувствительности.
В тест-наборы удобно вводить дифференциаторы, содержащие сразу несколько факторов устойчивости. Они входят и в набор Блэка (табл. 7). Расы с одним фактором вирулентности не способны заразить такие сорта. Расы с несколькими факторами вирулентности заражают их только в том случае, если факторы вирулентности комплементарны факторам устойчивости (например, раса 1.2 поражает дифференциатор R1R2, но не поражает дифференциатор R1R3). Раса со всеми факторами вирулентности поразит любые растения-дифференциаторы, входящие в набор. Применение сортов с несколькими факторами устойчивости позволяет уменьшить общее число растений-дифференциаторов в наборе.
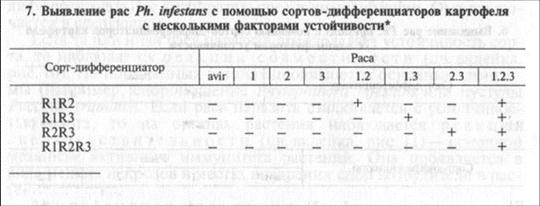
* Обозначения те же, что в таблице 6.
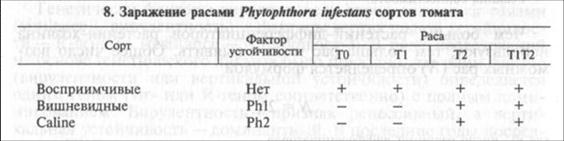
Помимо вирулентности к картофелю у Ph. infestans есть вирулентность и к томату. У патогена различают расы ТО, Т1 и Т2, а у томата — факторы устойчивости Phi и Ph2 (табл. 8). Раса ТО поражает картофель и восприимчивые сорта томата (их большинство), но не поражает сорта томата с факторами устойчивости. Раса Т1 поражает восприимчивые сорта томата а также вишневидные сорта (с небольшими ягодами) с фактором устойчивости Phi. Раса Т1Т2 поражает все сорта томатов. Хотя расы Т1 и Т2 были специализированы по отношению к томату, в последние годы обнаружили случаи поражения ими сортов картофеля.
При взаимодействии рас Puccinia graminis f. sp. tritici и пшеницы возможно больше реакций (табл. 9). В качестве дифференциаторов используют сорта пшеницы (Маркиз, Кубанка, Акме и др.), а также дикорастущие злаки.
Классификация рас возбудителя ржавчины пшеницы основана на группах реакций. Их три — устойчивая (У), восприимчивая (В), мезотетическая (М). Таким образом, число возможных рас (N) определяется формулой
N = 3 п,
где п — число растений-дифференциаторов.
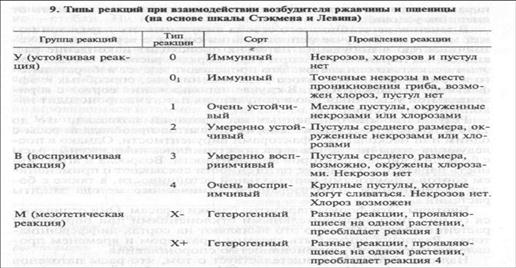
Основание степени равно трем, потому что имеется три типа реакции в ответ на заражение.
Определение рас возбудителя стеблевой ржавчины пшеницы обычно проводят в лабораторных условиях (температура 21 °С, освещенность около 10 ООО лк). Для поддержания влажности растения в вазонах покрывают специальными стеклянными колпаками. Изменение этих условий может несколько изменить тип реакции, например, привести к появлению мезотетических реакций. Для определения рас используют и полевой метод: высаживают дифференциаторы в полевых условиях и оценивают их пораженность.
На взаимоотношение рас и сортов могут влиять агрессивность паразитов и горизонтальная (полевая) устойчивость сортов. Так, падение агрессивности может помешать вирулентному паразиту поразить даже восприимчивый сорт растения-хозяина. Напротив, у авирулентного и высокоагрессивного паразита появляется некоторый шанс поразить устойчивый сорт. Высокая полевая устойчивость сорта несколько снижает вероятность его поражения комплементарной вирулентной расой. Даже авирулентная раса получит возможность поразить сорт с низкой полевой устойчивостью- Агрессивность и полевая устойчивость отражают жизнеспособность.
паразита и растения-хозяина соответственно и могут зависеть от внешних условий.
В течение вегетационного сезона вирулентность паразитов может меняться. В случае использования сортов с вертикальной устойчивостью в популяции патогена происходит накопление рас, способных поражать этот сорт. Разнообразие расового состава при этом несколько снижается. Это происходит за счет избирательного вегетативного и бесполого размножения рас, способных к эффективному паразитизму. В случае использования сортов с горизонтальной устойчивостью в популяциях патогена происходит накопление агрессивности.
Результаты многочисленных исследований показали, что до 1980-х годов в популяциях многих паразитов преобладали расы с одним или несколькими факторами вирулентности. Однако в последующие годы появились и распространились комплексные расы с большим числом факторов вирулентности. Возросла и агрессивность паразитов. Усиление патогенности связывают с применением сортов с факторами вертикальной устойчивости, а также с более интенсивным использованием химического метода защиты растений.
Биотипы. Эти формы паразитов близки к расам. Они отличаются от них лишь незначительными отклонениями при поражении растений-хозяев. Обычно это выявляют на сортах-дифференциаторах. Различия могут быть связаны с размером и временем проявления поражения, интенсивностью спороношения.
Наличие биотипов свидетельствует о том, что расы патогенов неоднородны по своему составу. Они могут быть представлены различными штаммами, отличающимися друг от друга своими физиологическими свойствами. Некоторые биотипы впоследствии могут стать новыми физиологическими расами.
Как указывалось ранее, паразиты с широкой специализацией способны поражать большое количество разнообразных видов растений-хозяев. Однако детальное изучение таких патогенов выявило генетическую неоднородность их популяций, различия в патогенности и вирулентности, а также физиологические особенности штаммов.
Один из наиболее изученных патогенов Botrytis cinerea — возбудитель серой гнили растений из семейств Розовых, Бобовых, Лилейных, Сложноцветных и др. При серой гнили органы растения поражаются в разной степени. В большей степени обычно пора жаются плоды и цветоносы, в меньшей степени листья, очень редко—стебли. На некоторых видах растений-хозяев обособились специализированные виды: Botrytis convallriae — на ландыше, Botrytis paeoniae — на пионе, Botrytis tulipae — на тюльпане. Эти виды реально представляют собой, интерстерильные группы (см. далее), обособленные друг от друга и от Botrytis cinerea. В пределах интерстерильных групп мицелии вегетативно совместимы между собой и образуют анастомозы и гетерокариоз. Такая обособленность позволяет им эффективнее поражать свои растения-хозяева. Эволюционно эти три вида, скорее всего, произошли от специализировавшихся штаммов Botrytis cinerea.
На интенсивность прорастания и эффективность заражения растений-хозяев может влиять инфекционная нагрузка (табл. 10).

Наименее специализированный вид Botrytis cinerea поражает широкий спектр растений-хозяев, но для эффективного заражения необходимо большое количество конидий. Остальные виды поражают преимущественно свой основной вид растения-хозяина, другие виды они поражают слабо. Однако при минимальной нагрузке спор только Botrytis convallariae способен поражать растения-хозяева. Это свидетельствует о наиболее узкой специализации Botrytis convallariae по сравнению с другими рассмотренными видами.
В целом, минимальные инфекционные нагрузки отличают облигатных паразитов с узкой специализацией от факультативных паразитов с широкой специализацией. Так, для успешного заражения возбудителю ржавчины при благоприятных условиях достаточно одной или нескольких урединиоспор, возбудителю фитофтороза картофеля — одной или нескольких зооспор или конидий (зооспорангиев). Для возбудителей серой гнили фузариозов споровые нагрузки значительно выше. Это связано с более отлаженным, более сбалансированным механизмом внедрения в растение- хозяина у облигатных паразитов по сравнению с факультативными. У последних заселение растений-хозяев определяется неоднородностью их химического состава, синтезом фитонцидов и фито-алексинов и способностью систем патогена снижать ингибирую-щее действие этих соединений.
Для вида Botrytis cinerea характерна клональная мозаичность популяций по вирулентности. Доминирующее положение в популяциях даже на одном виде растения-хозяина занимают клоны с широкой специализацией, поражающие довольно обширный спектр видов растений-хозяев. Другие клоны занимают промежуточное положение. Наконец третья группа клонов потенциально опасна только для определенных видов растений-хозяев. Различия в патогенное™, временной фактор и фаза развития растения-хозяина обусловливают проявление и доминирование определенных клонов возбудителя серой гнили.
Заражение нескольких видов растений-хозяев штаммами Botrytis cinerea выявило интересные закономерности (табл. 11).

Одни штаммы заражали все возможные растения-хозяева, другие — только некоторые. Часть штаммов были не способны заразить ни одно возможное растение-хозяин. Любопытно, что несмотря на то что все штаммы были выделены с земляники, некоторые из них не смогли перезаразить ее. Штаммы из второго варианта не заразили только землянику. Штаммы из большинства вариантов заражали растения-хозяева из различных семейств. Лишь штаммы из 10-го и 11-го вариантов заражали преимущественно растения из семейства Сложноцветных.
У разных растений-хозяев некрозы появлялись через разное время после заражения: на ландыше — только на 9-й день, у остальных—через 3...6 дней. Разница возникновения некрозов во времени не может не сказаться на продолжительности латентного периода (от заражения до новой генерации конидий). Это в итоге определяет численность клонов в популяции с определенными паразитическими свойствами.
Все эти факты свидетельствуют о слабой тенденции к сужению специализации отдельных штаммов. Однако она перекрывается сохраняющейся широкой специализацией у большинства штаммов паразита. Это сохраняет комплексную мозаичную структуру вида Botrytis cinerea в целом.
На эффективность заражения могут влиять климатические условия. Так, во влажных условиях некрозы образуются чаще, быстрее и они больших размеров. В засушливых условиях некрозы образуются реже и медленнее, они обычно небольшие.
Для Botrytis cinerea известна сезонная смена растений-хозяев. Ранней весной иногда поражаются мать-и-мачеха, одуванчик, хохлатка. Летом гриб часто инфицирует овощные и ягодные культуры. Осенью периодически поражаются цветочные культуры (астры; развитие патогена происходит преимущественно на цветоносах, соцветиях и цветках). Описаны случаи сапротрофного развития Botrytis cinerea на опаде.
Распределение растений-хозяев может оказывать определяющее влияние на распространение штаммов патогенов с широкой специализацией. Так, анализ развития серой гнили в условиях Московской области показывает, что отдельные растения-хозяева поражаются в значительной степени ежегодно. Растения-хозяева, на которых концентрируется основное количество возбудителя, А. Н. Лихачев назвал растениями-реципиентами. Для Botrytis cinerea растения-реципиенты — это земляника и малина (рис. 34). Благодаря преимущественному вегетативному размножению они формируют однородные популяции с выравненным по устойчивости фоном для патогена. На растениях-реципиентах происходит массовое накопление и сохранение инфекции в виде генетически различных клонов патогена, образующих центральные популяции — ядро вида Botrytis cinerea.
Первичное заражение растений-реципиентов осуществляется конидиями из их растительных остатков. Остальные растения-хозяева могут в значительной степени заражаться от растений-реципиентов. Это, например, пион, тюльпан, капуста, салат. Наконец растения, редко поражаемые серой гнилью (чеснок, лук-шалот, астра), могут получать инфекцию как от растений-реципиентов, так и от вторично поражаемых растений-хозяев. На всех этих растениях-хозяевах формируются краевые популяции Botrytis cinerea. В отдельных случаях возможны и обратные переходы штаммов из краевых популяций на растения-реципиенты.
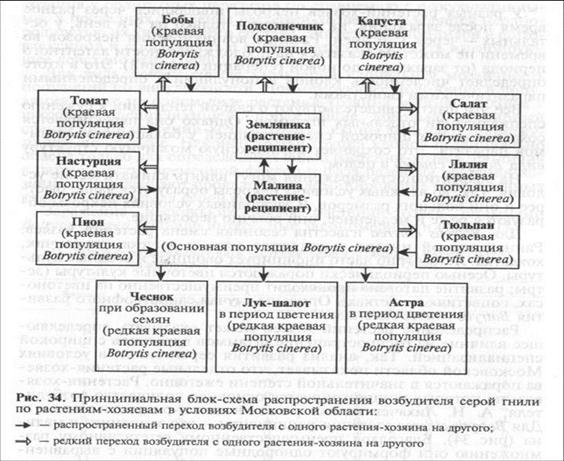
Случаи перезаражения вторично поражаемыми растениями друг друга серой гнилью очень редки. Это связано с некоторым сужением специализации штаммов Botrytis cinerea, которые на них паразитируют, а также с тем, что основная инфекция формируется на растениях-реципиентах.
Наличие выравненного по устойчивости фона хозяев-реципиентов создает предпосылки для возникновения популяций Botrytis cinerea со своей эволюционной судьбой и частично или полностью изолированных друг от друга за счет вегетативной несовместимости. Здесь уместно сравнить растения-реципиенты с промежуточными хозяевами возбудителя ржавчины. На растениях-реципиентах также происходит становление и распространение штаммов патогена, иногда даже более значительное, чем на промежуточных
хозяевах ржавчинных грибов. Однако на растениях-реципиентах не происходит половой гибридизации — самого главного механизма создания новых специализированных форм и рас, а также накапливания мутаций в рецессивном состоянии. Это не позволяет широко специализированным паразитам более эффективно эволюционировать в сторону повышения специализации.
Таким образом, массивы растений-реципиентов представляют собой опасность атаки широкоспециализированных паразитов как в агроценозах, так и в естественных сообществах. Возникающие на растениях-реципиентах и атакующие другие растения-хозяева группировки паразитов выполняют роль краевых популяций, осваивающих новых растений-хозяев. Даже неполная изоляция (на основе вегетативной несовместимости) снижает вероятность образования гетерокарионов между штаммами из центральных и краевых популяций паразита. Если гетерокарионы между ними все же возникают, то специализация по отношению к другим растениям-хозяевам снижается.
Многие виды растений-хозяев (такие, как земляника и малина) могут быть представлены как в биоценозах, так и в агроценозах. Поэтому природная инфекция широко специализированных паразитов на диких видах может оказать существенное влияние на структуру их популяций при выращивании растений-реципиентов и близкородственных растений в производственных условиях.