 2013-12-31
2013-12-31 1356
1356Большинству патогенов присуща как наследственная, так и модификационная изменчивость. Для ее создания у патогенов имеется несколько мощных механизмов. Их способность приобретать новые признаки и утрачивать прежние определяется растением-хозяином и внешней средой. Патогены способны очень оперативно изменяться при каких-либо внешних воздействиях на патосистему. Изменение агроэкологических факторов нередко приводило к смещениям нормы реакции патогенов, внедрение и размещение новых устойчивых сортов — к появлению новых рас.
Приведем основные механизмы и способы изменчивости и расообразования у грибных патогенов.
Половая гибридизация. Она связана с такими событиями в жизненном цикле патогенов, как мейоз и половой процесс. При мейозе происходит случайное распределение хромосом в дочерних ядрах, кроме того, нередок кроссинговер — обмен участками гомологичных хромосом. При половом процессе происходит слияние ядер (часто генетически разных), поэтому у дочерней формы сочетаются свойства родительских. В результате половой гибридиза ции происходит рекомбинация генетического материала паразитов и могут возникать их новые высокопатогенные формы.
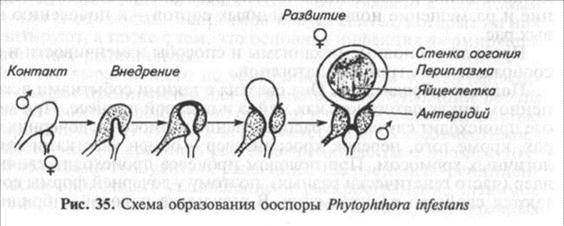
У Phytophthora infestans в результате полового процесса (слияния антеридия и яйцеклетки) образуются ооспоры (рис. 35). Мейоз осуществляется в антеридии и оогонии. При перекрестном скрещивании происходит контакт мицелиев двух типов спаривания — А1 и А2. Долгое время полагали, что штаммы А2 встречаются только в Мексике в высокогорной долине Толука, которую считали центром происхождения Phytophthora infestans. Поэтому полагали, что половая гибридизация возможна только там. В 1980— 2000-е годы А2 были найдены в большинстве регионов мира. Ооспоры обнаружили в некоторых странах Европы — России (Московская обл.), Польше, Германии, Голландии, странах Скандинавии. Как и в Мексике, в этих странах в природных популяциях Phytophthora infestans наблюдается очень высокое разнообразие расового состава.
Помимо контакта А1 и А2 ооспоры могут образовываться одним мицелием при самооплодотворении. Обычно это достигается его стимуляцией каким-либо химическим воздействием или внешним фактором. Однако возникшие таким образом ооспоры могут быть менее жизнеспособными и их вклад в генетическое разнообразие патогена ограничен.
На образование ооспор могут влиять экологические факторы, особенно относительная влажность воздуха. Образование ооспор затруднено при относительной влажности не < 80 %. При критическом понижении влажности оно прекращается. Это очень существенно, так как во многих полевых популяциях, где регулярно встречаются штаммы А1 и А2, образование ооспор может отсутствовать. В Мексике в высокогорной долине Толука относительная влажность воздуха и температурный режим оптимальны для развития возбудителя фитофтороза, что резко отличает ее от близлежащих регионов Мексики — пустынь на севере и тропических лесов на юге. Условия Толуки максимально способствуют образованию и прорастанию Ооспор. Повышение генетического разно-
образия паразита позволяет ему поражать многочисленных растений-хозяев (например, высокоустойчивый паслен Solanum demissum).
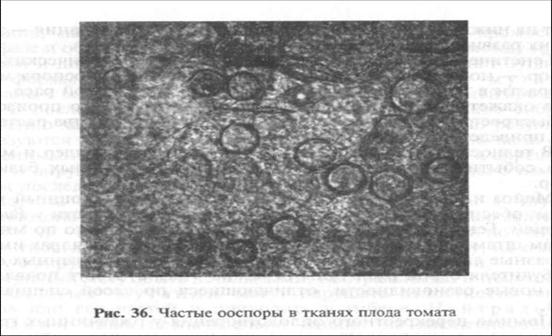
В Московской области ооспоры в органах растений картофеля развиваются довольно редко. В клубнях их образование затруднено. Однако в плодах томата при контакте А1 и А2 этот процесс протекает интенсивно. Плоды томата представляют собой удобный субстрат для формирования многочисленных ооспор (рис. 36). Более того, ооспоры Phytophthora infestans были обнаружены в семенах — на покрывающих их волосках и в теле зародыша. Это привело к неожиданно ранним вспышкам фитофтороза на рассаде томата. Но главное, такие ооспоры представляют реальную опасность для посадок картофеля, которые нередко располагают в непосредственной близости от посадок томата. Из этих ооспор могут сформироваться новые штаммы, патогенные для картофеля.
У возбудителя ржавчины Puccinia graminis половая гибридизация осуществляется на листьях промежуточных хозяев барбариса и ма-гонии, на которые попадают базидиоспоры (цв. вклейка, рис. 12). После их прорастания образуются половые структуры спермогонии. Они представляют собой небольшие пустулы с пучком улавливающих гиф. В спермогониях образуются спермации. При перекрестном оплодотворении они покидают один спермогоний и захватываются улавливающими гифами другого. Происходит плазмогамия. Образующийся дикариотический мицелий форми рует на нижней стороне листа более крупные образования — эции. В них развиваются эциоспоры.
Генетически каждая из образовавшихся дикариотических эци-оспор — новообразование. Попав на пшеницу, эциоспора может прорасти в урединиопустулу, соответствующую новой расе. Если раса окажется вирулентной для массива растений, то произойдет ее быстрое размножение и перезаражение большинства растений, что приведет к потере урожая.
В телиоспорах происходит слияние гаплоидных ядер и мейоз. Эти события предшествуют образованию гаплоидных базидио-спор.
Мейоз и половой процесс представляют собой мощный механизм, обеспечивающий высокий уровень изменчивости у Puccinia graminis. Генетическими методами было доказано, что по многим генам штаммы этого гриба гетерозиготны (в разных ядрах имеются разные аллели). При гибридизации специализированных форм возбудителя бурой ржавчины (Puccinia triticina) могут появляться его новые разновидности, отличающиеся по своей специализации.
Помимо перекрестного оплодотворения у ржавчинных грибов возможно и самооплодотворение, при котором спермации и улавливающие гифы принадлежат одному штамму. Самооплодотворение также способствует расообразованию. Так, при самооплодотворении из одной физиологической расы возбудителя карликовой ржавчины ячменя (Puccinia anomala) было получено шесть физиологических рас. Сходные результаты были получены и для возбудителя корончатой ржавчины овса (Puccinia coronifera).
В случае отсутствия промежуточных хозяев спермогониев и эциев в цикле развития Puccinia graminis не будет. Доказано, что в этом случае интенсивность образования рас существенно снижается. Кроме того, у некоторых популяций Puccinia graminis и других ржавчинных грибов может выпадать из цикла развития и тели-опустула. В популяциях, представленных только урединиопусту-лами, разнообразие рас будет ограниченным.
Сходным образом образование новых рас происходит и у других паразитов — головневых и мучнисторосяных грибов, а также возбудителя парши яблони аскомицета Venturia inaequalis. Телиос-поры головневых и аскоспоры возбудителей мучнистых рос и парши яблони, образованию которых предшествовали мейоз и половой процесс, способствуют образованию новых рас.
Мутации. У паразитов мутации могут быть спонтанными (не вызванными внешними факторами) и индуцированными (вызванными внешними факторами). Мутации вызываются действием химических веществ (нитрозометилмочевина и этилме-тансульфонат), а также ультрафиолетового облучения.
У грибов различают геномные, хромосомные и генные мутации. Геномные мутации связаны с нарушением процесса деления, например, нерасхождение обоих наборов хромосом в анафазе и объединение их в одном ядре, кариогамия между гаплоидным и диплоидным ядрами. В результате возникают эуплоид-ные формы (Зл, 4л, 5л и выше). Хромосомные мутации связаны с выпадением хромосомы или добавкой лишней хромосомы. В последнем случае образуются трисомики. Такие события особенно характерны при нарушениях хода мейоза. В результате образуются нестабильные анеуплоидные формы, очень часто с измененными свойствами (в том числе по способности к скрещиванию и вирулентности). Генные мутации связаны с изменением последовательности нуклеотидов в ДНК.
Частота мутаций зависит от вида гриба и гена. У Erysiphe graminis она не превышает 2 • 10~8 %, у Helminthosporium sativum — 5 • Ю-2...5 • 10_3 %. У Pyricularia oryzae частота мутаций по гену вирулентности составляла 1,2 • Ю-1 %, а по другому гену, не связанному с вирулентностью, мутации обнаружены не были.
Мутации могут быть летальными, нейтральными и полезными. Летальные мутации могут нарушать целостность генома гриба или его частей и приводить к гибели. Нейтральные мутации лишь незначительно влияют на жизнеспособность гриба. Полезные мутации приводят к расширению вирулентности. Например, мутанты возбудителя бурой ржавчины пшеницы гриба Puccinia recondita расширили свою вирулентность в 50 раз по сравнению с мутантами, понизившими ее. Мутации могут сохраняться в популяции в рецессивном или гетерокариотическом состоянии.
Мутации приводят к формированию новых рас. Этот способ имеет особенно большое значение у паразитов, не имеющих в своем цикле половой стадии, а следовательно, и половой гибридизации.
Помимо вирулентности мутации могут изменять морфологические свойства мицелия, его отношение к температуре, а также приводить к повышению устойчивости к фунгицидам и антибиотикам.
В ряде случаев мутанты с расширенной вирулентностью и повышенной устойчивостью к фунгицидам могут понижать свою агрессивность и жизнеспособность. Однако они могут закрепляться в популяции в рецессивном или гетерокариотическом состоянии. Со временем жизнеспособность мутантов может повыситься и тогда благодаря бесполому размножению их численность в популяции патогена может резко возрасти. Далее последует потеря устойчивости сорта или эффективности химического препарата. Такие случаи широко распространены в сельскохозяйственной практике.
Вегетативная совместимость / несовместимость. Гетероплазмоз. Гетерокариоз. Еще один довольно распространенный механизм изменчивости и расообразования — образование контакта (пере мычка между гифами — так называемый анастомоз) между клетками различных мицелиев с последующим слиянием их цитоплазм. Такой контакт между клетками возможен только при условии вегетативной совместимости мицелиев. Контактирующие поверхности гиф сначала слипаются, затем происходит ферментативное разрушение клеточной стенки в месте контакта. В результате образуется мицелий с единой смешанной цитоплазмой — гетероплазмон (от гр. heteros — другой и plasma — образование), в которой находятся и функционируют ядра из исходно различных мицелиев — гетерокарион (отгр. heteros — другой и karyon — ядро).
Гетерокарион может образовываться и в том случае, если в части ядер мицелия произошли мутации.
Вегетативная совместимость известна у многих грибов: Aspergillus, Botrytis, Ceratocystis, Cochliobolus, Cryphonectria, Diaporthe, Fusarium, Hypochnus, Hypoxylon, Penicillium, Phytophthora, Podospora, Sclerotinia, Septoria, Trichoderma, Typhula и Verticillium. Система вегетативной совместимости / несовместимости находится под жестким генетическим контролем и может ограничивать образование анастомозов, слияние цитоплазм, а также гибель гетерокариотич-ной клетки из-за Несовместимости различных ядер. Вегетативно совместимые штаммы образуют группы вегетативной совместимости.
Если вегетативная несовместимость работает на стадии анастомозов, то группы вегетативной совместимости называют а н а с -томозными группами (АГ) или интерстерильными группами. Анастомозные группы описаны у базидиомицета Hypochnus solani (анаморфа дейтеромицет Rhizoctonia solani). Их известно 10 —АП, АГ2, АГЗ,..., АГ9, АПО. Первые четыре АГ-группы встречаются повсеместно, остальные — в отдельных регионах мира. Анастомозные группы характеризуются различной степенью изолированности друг от друга. Кроме того, они имеют различную способность образовывать базидиальную стадию. Так, АП и АГ4 легко и регулярно формируют базидиальную стадию, а АГ2 и АГЗ — нет. Анастомозные группы отличаются друг от друга по особенностям паразитизма и специализации к растениям-хозяевам, что обеспечивает сложную мозаичную структуру комплексного вида Rhizoctonia solani (табл. 12).
12. Встречаемость анастомозных групп Rhizoctonia solani на различных растениях-хозяевах

У возбудителя тифулеза озимых злаков Typhula ishikariensis описано три разновидности: var. ishikariensis, var. idahoensis и var. canadensis. В их пределах на основе морфологических и экологических отличий выделено несколько биотипов. Штаммы этих разновидностей и биотипов иногда образуют анастомозы между собой, иногда — нет. Поэтому они представляют собой интерстерильные группы, аналогичные анастомозным группам Rhizoctonia solani.
Интерстерильные группы были открыты у другого комплексного вида — Armillariella mellea (опенка осеннего), поражающего хвойные, лиственные и плодовые породы деревьев. Они отличались между собой специализацией по растениям-хозяевам, составу ферментов и последовательностям ДНК.
В случае успешного контакта гиф с образованием анастомоза может сказаться несовместимость между цитоплазмами. Произойдет их взаимное отторжение, сопровождающееся разрушением органелл, в результате мицелий погибнет.
В случае совместимости цитоплазм может сказаться последний фактор — несовместимость4 ядер. В несептированных мицелиях хитридиомицетов, оомицетов и зигомицетов возможно равномерное перемешивание ядер и стабильное существование сбалансированных гетерокарионов со случайным соотношением ядер. Миграция ядер осуществляется благодаря микротрубочкам цито-скелета. У аскомицетов, базидиомицетов и дейтеромицетов свободная миграция ядер ограничивается септами. Однако ограниченная миграция ядер все же возможна через центральную пору. У одних грибов (Botrytis, Helminthosporium) соотношение ядер стабильно и не зависит от адаптивных свойств находящихся в них генов, у других (Verticillium, Pyricularia, Fusarium) — гетерокарионы образуются только в районе контакта анастомозирующих мицелиев. Поскольку ядра не мигрируют к центру колоний, гифы там преимущественно гомокариотичны.
Установлено, что конидии многих дейтеромицетов гетерокари-отичны.
В случае несовместимости ядер или отдельных генов и их продуктов происходит либо возврат к гомокариону, либо нарушение нормального физиологического состояния мицелия и его гибель. Это происходит вследствие конкуренции между ядрами (конкурентной экспрессии их генов и несовместимости посттрансляционных продуктов). Другая возможная причина этих явлений — нарушение обмена веществ и неспособность гетерокариотического мицелия эффективно использовать субстрат.
Случаи образования новых рас при вегетативной совместимости описаны исследовательницей Малколмсон для Phytophthora infestans. Она использовала суспензии конидий рас 1.2.3.7 и 4. Исходные конидии были смешаны и помещены на листья сорта R1R2R3R4. После заражения на листьях появилось слабое споро ношение. Из него приготовили новую суспензию и вновь заразили сорт R1R2R3R4. На листьях вновь появилось спороношение. Эту процедуру повторили несколько раз. При последнем цикле заразили не только R1R2R3R4, но и R7. В обоих случаях заражение прошло успешно. В других случаях после нескольких циклов наблюдали потерю одного из факторов вирулентности. Эти эксперименты доказали вегетативную гибридизацию у Phytophthora infestans, ведущую к образованию гетерокарионов по факторам (генам) вирулентности и образованию новых рас.
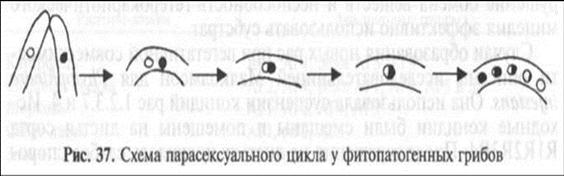
Парасексуальный цикл. Первый этап цикла может включать реакцию совместимости: контакт мицелиев, образование между ними анастомозов, а затем гетерокариона и гетероплаз-мона (рис. 37).
Далее происходит парасексуальный процесс — кариогамия, слияние ядер гетерокариона. Это редкое событие, его вероятность составляет порядка 10~6. В результате возникают генетически смешанные ядра с увеличенной плоидностью. При слиянии гаплоидных ядер образуется диплоидное ядро (так называемый гетерозиготный диплоид). При слиянии гаплоидных и диплоидных ядер образуется триплоидное ядро, диплоидных ядер — тетрапло-идное ядро. Нередко новообразованные ядра представляют собой несбалансированные хромосомные системы.
Парасексуальный процесс может происходить и с участием го-мокариотических ядер.
Митотические деления новых смешанных ядер происходят нередко с нарушениями. Часто они сопровождаются гаплоиди-з а ц и е й — уменьшением плоидности ядер, очень часто возвратом к их исходной плоидности. Этот процесс сопровождается потерей лишних наборов хромосом или отдельных хромосом. В результате многие ядра полностью возвращаются к исходному статусу. Но в некоторых ядрах происходит митотическая рекомбинация. Возникают так называемые сегреганты (от англ. segregation — расщепление). Они в большей или меньшей степени сочетают в себе хромосомы и гены обоих исходных ядер. Стабильность таких новообразованных ядер различна. Некоторые генетически нестабильны и гибнут, другие, напротив, делятся интенсивнее других. У фитопатогенов таким образом могут возникать ядра-сегреганты, сочетающие рецессивные гены вирулентности исходных ядер. Это позволяет рецессивным генам оказывать определяющее влияние на
фенотип (из авирулентного он становится вирулентным), а также расширять вирулентность — увеличивать число генов вирулентности. Таким способом могут образовываться генетически стабильные комплексные расы фитопатогенных грибов.
Мобильные генетические элементы — транспозоны. У многих грибов обнаружены мобильные лбкусы, способные менять свое положение в геноме, перемещаясь по одной или даже по разным хромосомам. В них могут вовлекаться многие важные гены, в том числе и гены вирулентности. Когда эти гены попадают в новое место, может сработать так называемый эффект положения. Он проявляется в том, что функционирование генов в новом месте изменяется. Это приводит к появлению новых (нередко нестабильных) свойств патогена.
Транспозоны фитопатогенных грибов напоминают транспозоны одного из наиболее изученных генетических объектов — дрозофилы. Доказано, что мутации некоторых генов Fusarium oxysporum связаны со вставкой в них транспозона типа Tc-mariner. Такие мутации изменяют физиологические свойства гриба.
Высказано предположение, что по своим свойствам транспозоны близки к вирусам на этапе их встраивания в геном хозяина.
Дрейф генов. Это случайные изменения частот генов, обычно происходящие в течение вегетационного сезона и могут быть вызваны перенесением неблагоприятных условий. Такие изменения могут происходить при изменении численности популяций патогенов. Так, ее уменьшение наблюдается поздней осенью в конце вегетационного сезона и после проведения защитных мероприятий (севооборотов и применения фунгицидов). Увеличение популяций патогенов может происходить, когда они попадают на массивы восприимчивых растений (к примеру, на растения-реципиенты). Однако массово размножиться могут только некоторые генотипы паразитов, которые случайно первыми попали на растения. Такую возможность называ-ютэффектом бутылочного горлышка.
В результате дрейфа генов распределение рас паразитов в первичных очагах инфекции может иметь случайный (мозаичный) характер. Но далее происходит накопление патогенных и хорошо приспособленных форм паразита. Это происходит вследствие действия естественного отбора. Так, у Phytophthora infestans в полевых популяциях в первой половине вегетационного сезона обычно встречаются расы с разным количеством факторов вирулентности. Однако к концу сезона преобладают комплексные расы, включающие большинство или все факторы вирулентности.
Изменения генетического состава популяций могут быть обусловлены действием дрейфа генов и отбором. Дрейф генов может приводить к резким различиям между популяциями фитопатогенов во времени и в пространстве.
Миграции. В перемещении патогенных организмов между континентами, странами и внутри них может принимать участие хо зяйственная деятельность людей, воздушные течения, переносчики (Горленко, 1975). Многие патогены и их формы пересекали значительные расстояния с посадочным материалом.
Межконтинентальные миграции происходят при содействии человека с грузоперевозками. Так, из Мексики через США в Европу мигрировала Phytophthora infestans при перевозке клубней картофеля. Первая миграция была осуществлена в 1840-е годы. Точно известно, что в результате нее в Бельгии, Ирландии, и Великобритании появились штаммы А1. Они вызвали опустошительные эпифитотии на картофеле, приведшие к тяжелым экономическим и политическим последствиям. Вторая миграция была осуществлена в 1970—1980-е годы. Из Мексики были поставки картофеля в Европу. В этот же период из соседней с Мексикой Кубы картофель регулярно поставляли в регионы бывшего СССР. При второй миграции в Европе появились и распространились штаммы не только А1, но и А2. Эти мигранты характеризовались новыми генетическими свойствами, большей патогенностью, а также повышенной устойчивостью к фунгицидам. Это позволило им значительно потеснить штаммы «старого типа». Наличие обоих типов спаривания (А1 и А2) во многих странах привело к возможности в них половой гибридизации — мощного источника расширения вирулентности и появления новых рас.
Межконтинентальные миграции до сих пор представляют опасность для тех патогенов, которые уже распространены в России и других странах Европы, но центры их происхождения находятся на других континентах. Для примера можно привести Xanthomonas citri (возбудитель бактериального рака цитрусовых), который был завезен из Китая в США.
В последние годы возможны и обратные примеры, когда из мест интенсивной интродукции и искусственного выведения сортов происходит распространение возбудителей болезней. Так, в Америку из стран Европы попали рак и кольцевая гниль картофеля.
Внутриконтинентальные миграции осуществляются с воздушными течениями, переносчиками, посевным и посадочным материалом. Возможен комбинированный путь расширения ареала патогенов — занос сначала с семенами, а затем распространение по воздуху. Таким способом распространился и продолжает свое шествие по Европе возбудитель белой ржавчины хризантем (Puccinia horiana). Перенос ветром урединиоспор на расстояния около 1 км осуществляется очень быстро.
Распространение возбудителя бактериального ожога плодовых (Erwinia amylovora) шло несколько медленнее. Первичный очаг был выявлен в Англии в 1957 г. Только к 1970 г. в Англии и странах Скандинавии были зарегистрированы вторичные очаги болезни. Распространение источников осуществлялось перелетными птицами, в основном скворцами.
Миграции внутри страны осуществляются так же, как и внутриконтинентальные миграции. Они тесно связаны с хозяйственной деятельностью человека той или иной страны и для каждой из них могут быть специфичны. К примеру, в Российской Федерации расширение площадей возделывания кукурузы на север в 1960—1970-е годы способствовало продвижению туда узкоспециализированного возбудителя головни кукурузы (Ustillago zeae), а также широкоспециализированных возбудителей фузари-оза (Fusarium spp.). Большое значение в условиях России имеет распространение конидий возбудителя фитофтороза и урединиоспор возбудителя ржавчины посредством движения воздушных масс в атмосфере. В США выявлен специальный коридор (Puccinia path) потоков урединиоспор с севера на юг и с юга на север страны.
Распространение патогенов возможно путем интродукции растений в парковых зонах и ботанических садах.
Внутри- и межхозяйственные миграции осуществляются между отдельными хозяйствами или полями севооборотов одного и того же хозяйства. Внутрихозяйственные миграции могут происходить после заноса какого-либо патогена из других мест. Этот тип миграций очень характерен для возбудителей ложных и настоящих мучнистых рос.
Миграции из природных очагов представляют собой занос инфекционного начала с дикорастущих растений (где патогены могут перезимовывать) на культурные растения. Например, культурные крестоцветные ежегодно могут заражаться белой ржавчиной. Ее возбудитель — широкоспециализированный паразит оомицет Albugo Candida перемещается на них с сорных крестоцветных (ярутка, пастушья сумка). Большую роль в таких миграциях могут играть растения-реципиенты, произрастающие в агроценозах.
Переходящие в агроценозы фитопатогены нередко эволюционируют в пределах агроценоза независимо от исходных форм из биоценозов. Они обособляются от этих исходных форм, образуя специализированные формы у облигатных паразитов, интерстерильные группы или группы вегетативной несовместимости. Образование таких групп особенно характерно для видов родов Botrytis, Rhizoctonia, Armillariella.
Наличие всех этих типов миграций обусловливает появление уже сформировавшихся новых видов патогенов, а также их специализированных форм, рас, штаммов. В случае успешной адаптации они могут представлять огромную опасность для местных видов растений-хозяев, у которых нет против них ни иммунитета, ни, возможно, механизмов его приобретения.








