 2013-12-31
2013-12-31 852
852Хлоропласты и их элементарная структура.
Структурная организация фотосинтетического аппарата.
Фотосинтез как процесс трансформации энергии света в энергию химических связей.
Общее уравнение фотосинтеза, его компоненты.
Сущность и экологическое значение фотосинтеза.
История развития учения о фотосинтезе.
Фосфорилирование. Электрохимический потенциал - основа и движущая сила фосфорилирования.
Строение и свойства энергетических молекул (АТФ, НАДФ). Образование макроэргических соединений в процессе окислительно-восстановительных реакций.
Автотрофность и гетеротрофность. Однообразие элементарных энергетических процессов у различных организмов.
Источники энергии в биологических системах.
Функциональное взаимодействие различных органоидов клетки.
Особенности строения клеточных органоидов в связи с их биологическими функциями.
Основные структурные элементы растительной клетки.
Растительная клетка как организм и как элементарная структура многоклеточного растительного организма.
Космическая роль зеленого растения.
Разнообразие организмов, характеризующихся фототрофным образом жизни. Основные признаки, определяющие их единство.
Экологический и эволюционный аспекты физиологии растений.3. Связь физиологии растений с другими биологическими науками - биохимией, биофизикой, молекулярной биологией, генетикой и др. 4. Методологические аспекты современной физиологии, как подходы к изучению живых систем на разных уровнях исследования (клеточный, организменный, уровень биоценоза и биосферный).
Лебедев. «Физиология растений». Дежавю. стр.3
«Физиология растений». Дежавю. стр. 117
В тетради.
В тетради
«Физиология растений». Дежавю. стр.277
«Физиология растений». Дежавю. стр. 156
«Физиология растений». Дежавю. стр.59
«Физиология растений». Дежавю. стр.106
«Физиология растений» Дежавю. стр. 59
«Жизнь зеленого растения». Дежавю. стр.113
Хлорофиллы
У всех высших растений, водорослей и цианобактерий содержится хлорофилл a, хлорофилл b имеется у высших растений и зеленых водорослей. Хлорофилл с, лишенный фитола, содержится в бурых и диатомовых водорослях, хлорофилл d – в красных водорослях. Фотосинтезирующие зеленые бактерии имеют бактериохлорофиллы c и d, пурпурные бактерии - бактериохлорофиллы a и b.
У хлорофилла а четыре пиррольных кольца соединены между собой метиновыми мостиками (=СН-), образуя порфириновое кольцо. Кроме того, атомы азота пиррольных колец связаны с атомом магния. С порфириновым ядром соединено циклопентановое кольцо, образованное остатком кетопропионовой кислоты и содержащее активные карбонильную (С=О) и метилированную карбоксильную (О=С-О-СН3) группы. Структура, состоящая из порфиринового ядра и циклопентанового кольца, называется форбином. Боковая цепь, состоящая из пропионовой кислоты и непредельного спирта фитола, связана с атомом углерода IV пиррольного кольца (рис. 5.1). Хлорофилл, лишенный фитола, называется хлорофиллидом. Если атом магния замещен протоном, то такое соединение носит название феофитина. Активность хлорофиллов, также как и других пигментов, обусловлена наличием большого количества двойных связей с делокализованными электронами.
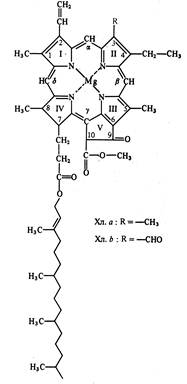
Рис. 5.1. Структурные формулы хлорофиллов a и b (по В. В. Полевому).
Хлорофиллы хорошо растворимы в органических растворителях (этиловом эфире, бензоле, хлороформе, ацетоне, этиловом спирте) и нерастворимы в воде. Хлорофиллы имеют максимумы поглощения света в красной и синей частях спектра. Растворы хлорофиллов обладают флуоресценцией и фосфоресценцией.
5.1.2. Каротиноиды
Каротиноиды – жирорастворимые пигменты, присутствующие в хлоропластах всех растений. Они входят в состав хромопластов в незеленых частях растений, например, корнеплодов моркови. К каротиноидам относят 3 группы соединений: 1) оранжевые или красные каротины, 2) желтые ксантофиллы, 3) каротиноидные кислоты. Каротины и ксантофиллы состоят из 8 остатков изопрена, которые образуют цепь конъюгированных двойных связей (рис. 5.2). Основные каротиноиды - b-каротин, лютеин, виолаксантин и неоксантин.
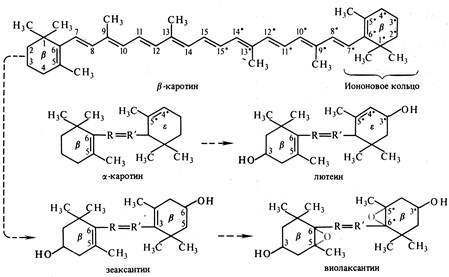
Рис. 5.2. Структурные формулы каротиноидов и последовательность их превращений (по В. В. Полевому).
Каротины и ксантофиллы растворимы в хлороформе, бензоле, сероуглероде, ацетоне. Каротины хорошо растворяются в эфирах, но плохо в спиртах, а ксантофиллы наоборот. Каротиноиды имеют максимумы поглощения в фиолетово-синей и синей частях спектра света. Они не способны к флуоресценции.
Главные функции каротиноидов: поглощение света в качестве дополнительных пигментов, защита молекул хлорофиллов от необратимого фотоокисления, тушение активных радикалов, участие в фототропизме, так как способствуют определению направления роста побега.
5.1.3. Фикобилины
Сине-зеленые и красные водоросли помимо хлорофилла а и каротиноидов содержат пигменты фикобилины. Их молекула состоит из 4 последовательных пиррольных колец (рис. 5.3). Фикобилины являются хромофорными группами глобулиновых белков фикобилипротеинов. Они делятся на 3 группы: 1) фикоэритрины – белки красного цвета, 2) фикоцианины – сине-голубые белки и 3) аллофикоцианины – синие белки. Все они обладают флуоресценцией и растворимы в воде.
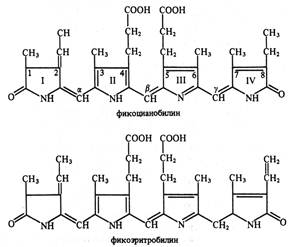
Рис. 5.3. Структурные формулы фикобилинов (по В. В. Полевому).
Фикобилины имеют максимумы поглощения в оранжевой, желтой и зеленой частях спектра света. Это позволяет водорослям полнее использовать свет, проникающий в воду. Вода обладает светопоглощающей способностью. На глубине около 30 м полностью исчезают красные лучи, около 180 м – желтые, 320 м – зеленые, а на глубину более 500 м не проникают синие и фиолетовые лучи. Фикобилины – это дополнительные пигменты, участвующие в светособирающем комплексе. Около 90 % энергии света, поглощенного фикобилинами, передается на хлорофилл а.
У растений имеется фикобилин фитохром. Он не участвует в фотосинтезе, но является фоторецептором красного и дальнего красного света и выполняет регуляторные функции в клетках растений.








