 2013-12-31
2013-12-31 920
920Цикл Кребса.
Пути окисления органических веществ. Основные пути диссимиляции углеводов.
«Физиология растений». Дежавю. стр. 130
Цикл ди- и трикарбоновых кислот (цикл Кребса)
Аэробная фаза дыхания локализована в митохондриях. Пировиноградная кислота окисляется до воды и углекислого газа в дыхательном цикле, получившем название цикла ди- и трикарбоновых кислот или цикла Кребса в честь английского биохимика Г. Кребса, описавшего этот путь (рис 6.2.). В этом цикле окисляется не сама пировиноградная кислота, а ее производное – ацетилкоэнзим А. Он образуется в результате окислительного декарбоксилирования пировиноградной кислоты. Процесс этот состоит из ряда реакций и катализируется сложной мультиферментной системой, состоящей из трех ферментов и пяти коферментов, и названной пируваткарбоксилазой.
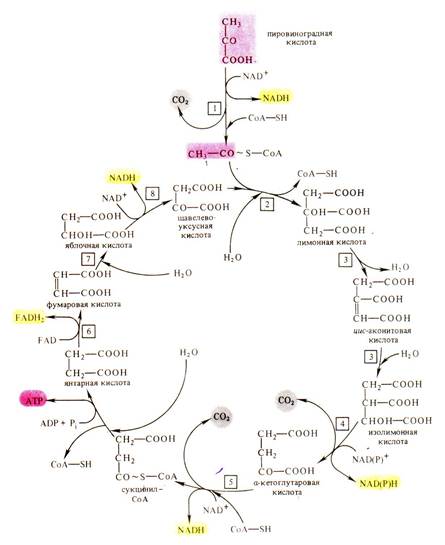
Рис. 6.2. Цикл Кребса (цикл ди- и трикарбоновых кислот).
1 – мультиэнзимный комплекс окислительного декарбоксилирования пировиноградной кислоты, 2 – цитратсинтаза, 3 – аконитатгидратаза, 4 – изоцитратдегидрогеназа, 5 – мультиэнзимный комплекс окислительного декарбоксилирования α-кетоглутаровой кислоты, 6 – сукцинатдегидрогеназа, 7 – фумаратгидратаза, 8 – малатдегидрогеназа (по В. В. Полевому).
|
|
|
При окислении одной молекулы пировиноградной кислоты образуется 3 молекулы НАДН, 1 молекула НАДФН и 1 молекула ФАДН2, при окислении которых в дыхательной электронтранспортной цепи синтезируется 14 молекул АТФ. Кроме того, 1 молекула АТФ образуется в результате субстратного фосфорилирования.
Дыхательная электронтранспортная цепь
и окислительное фосфорилирование
Дыхательная электронтранспортная цепь состоит из переносчиков электронов, которые передают электроны от субстратов на кислород. Расположение переносчиков определяется величиной их окислительно-восстановительного потенциала. Цепь начинается с НАДН, имеющего потенциал –0,32 В, и кончается кислородом с потенциалом +0,82 В. Переносчики расположены по обеим сторонам внутренней мембраны митохондрий и пересекают ее. На внутренней стороне мембраны, расположенной к матриксу митохондрии, два протона и два электрона от НАДН переходят на флавинмононуклеотид и железосерные белки. Флавинмононуклеотид, получив протоны, восстанавливается и переносит их на внешнюю сторону мембраны, где отдает протоны в межмембранное пространство. Железосерные белки, находящиеся внутри мембраны, передают электроны от НАДН окисленному убихинону Q. Он, присоединив еще два протона, диффундирует в мембране к цитохромам. Цитохром b 560 отдает два электрона убихинону, который, присоединив еще два протона из матрикса, передает два электрона цитохрому b 556 и два электрона цитохрому c 1, а протоны выходят в межмембранное пространство. На наружной стороне мембраны цитохром с, получив два электрона от цитохрома c 1, передает их цитохрому а, который переносит их через мембрану на цитохром а 3. Цитохром а 3, связывая кислород, отдает ему электроны. Кислород присоединяет два протона с образованием воды (рис. 6.5).
|
|
|
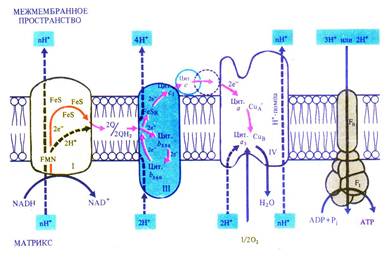
Рис. 6.5. Локализация электрон- и протонтранспортных реакций во внутренней мембране митохондрий (по В. В. Полевому).
Таким образом, транспорт электронов в дыхательной электронтранспортной цепи сопровождается трансмембранным переносом протонов. Возникающая разность потенциалов по обеим сторонам внутренней мембране митохондрий используется для синтеза АТФ (окислительное фосфорилирование), как это было показано в разделе 5.2.2. В результате прохождения двух электронов по цепи образуется 3 молекулы АТФ.








