 2014-01-25
2014-01-25 1123
1123Мембрана клетки состоит из двойного слоя липидов, между которыми находятся молекулы белка (Рис.8). Часть белков находится на внутренней или наружной поверхности мембраны, а часть пронизывает ее насквозь (интегральные белки). В некоторых интегральных белках есть поры (ионные каналы), через которые проходят ионы, участвующие в образовании мембранных потенциалов.
В возникновении и поддержании мембранного потенциала покоя принимает участие два основных белка:
· Белок, образующий Na+/К+ - насос, который за счет энергии АТФ активно перекачивает Na+ из клетки наружу, а К+ внутрь. В результате концентрация К+ становится внутри клетки выше, а снаружи оказывается больше Na+.
· Белок, который служит каналом утечки К+, через который ионы К+ в силу диффузии стремятся выйти из клетки, где они в избытке.
В результате работы двух вышеуказанных каналов мембрана снаружи оказывается заряжена положительно, а внутри отрицательно (Рис. 9). То есть мембрана в состоянии покоя поляризована и имеет разность потенциалов по обе стороны, которая называется потенциал покоя, равный примерно минус 70 мВ (для нейрона), а для мышечного волокна минус 90 мВ.
В основе возбуждения нервных и мышечных клеток лежит повышение проницаемости мембраны для ионов Na+ (открытие Na+ - каналов). Внешнее раздражение вызывает перемещение заряженных частиц внутри мембраны и снижение исходной разности потенциалов (деполяризация мембраны). Небольшая деполяризация приводит к локальным изменениям функционирования клетки (подпороговая реакция).
При увеличении раздражения изменение мембранного потенциала достигает порога возбудимости (критического уровня деполяризации – около 20 мВ). При этом величина потенциала покоя снижается до минус 50 мВ.
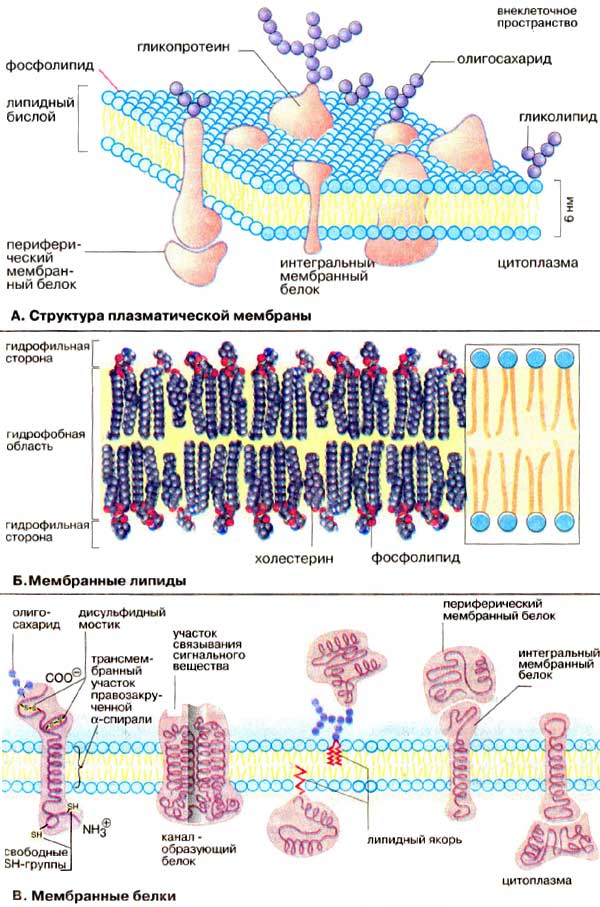
Рис.8. Структура биомембран.
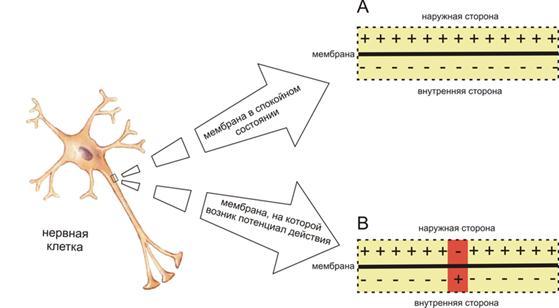
Рис.9. Схема распределения зарядов по разные стороны мембраны возбудимой клетки в спокойном состоянии (A) и при возникновении потенциала действия (B).
В результате открывается большинство Na+ - каналов, после чего резко ионы Na+ входят внутрь клетки, изменяя мембранный потенциал, что регистрируется в виде потенциала действия (ПД). В месте возбуждения внутренняя сторона меняет заряд на положительный, а внешняя – на отрицательный (Рис.9).
Процесс открывания Na+ - каналов кратковременный (1-2 мс), после чего Na+ - каналы закрываются. К этому моменту достигает большой величины нараставшая при возбуждении проницаемость К+ - каналов. Выходящие из клетки ионы К+ вызывают быстрое снижение потенциала действия и происходит окончательное восстановление исходного заряда. Поэтому в ПД различают кратковременную высоковольтную часть – пик (спайк) и длительные малые колебания – следовые потенциалы.
В процессе восстановления после ПД Na+/К+ - насос обеспечивает «откачку» излишних ионов Na+ наружу и «накачивание» потерянных ионов К+ внутрь. То есть возвращение исходной асимметрии их концентрации по обе стороны мембраны. На работу этого механизма тратится около 70% всй необходимой клетке энергии.
Возникновение возбуждения (ПД) возможно при сохранении достаточного количества Na+ в окружающей клетку среде. Потери Na+ организмом (при длительной мышечной работе с потом) нарушают нормальную деятельность нервных и мышечных клеток, снижают работоспособность.
При недостатке кислорода (кислородный долг при мышечной работе) процесс возбуждения нарушается из-за инактивации вхождения Na+ в клетку и клетка становится невозбудимой. На процесс инактивации натриевого механизма влияет концентрация ионов Са2+ в крови. При повышении содержания Са2+ снижается клеточная возбудимость, а при его недостатке возбудимость повышается и появляются непроизвольные мышечные судороги.








