 2014-01-25
2014-01-25 1668
1668В нормальных физиологических условиях глубина вдоха может быть ограничена только физическими свойствами легочной ткани и грудной клетки. Сопротивление раздуванию легких, которое возникает при поступлении в них воздуха, обусловлено растяжимостью их соединительной ткани и сопротивлением дыхательных путей потоку воздуха. Мерой эластических свойств легочной ткани является растяжимость легких, которая характеризует степень увеличения объема легких в зависимости от степени уменьшения внутриплеврального давления:
С = dV/dР,
где С — растяжимость (англ. — compliance), dV — изменение легочного объема (мл), а dР — изменение внутриплеврального давления (см водн. ст.). Растяжимость характеризует количественно степень изменения объема легких у человека в зависимости от степени изменения при вдохе внутриплеврального давления. Грудная клетка также обладает эластическими свойствами, поэтому растяжимость тканей легких и тканей грудной клетки обусловливает эластические свойства всего аппарата внешнего дыхания человека.
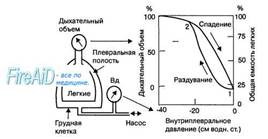
Рис. 10.6. Кривая растяжимости (комплайенса) легких. Кривые справа показывают изменение дыхательного объема легких и общей емкости легких, возникающие при изменениях внутриплеврального давления без учета влияния тканей грудной клетки. Легкие полностью не спадаются, если внутриплевральное давление становится равным нулю (точка 1). Кривые комплайенса совпадают в точке 2 при большом объеме в легких, когда легочная ткань достигает предела эластического растяжения. Вд — внутриплевральное давление. Слева — схема регистрации изменений внугриплеврального давления и дыхательного объема легких.
На рис. 10.6 показано изменение легочного объема, которое возникает при изменениях внутриплеврального давления. Восходящая и нисходящая линии характеризуют раздувание и спадение легких соответственно. Фрагмент рис. 10.6 слева демонстрирует, каким образом могут быть измерены значения легочного объема и внутриплеврального давления, вынесенные на график. Объем легких не уменьшается до нуля, когда величина внутриплеврального давления становится равной нулю. Раздувание легких с уровня их минимального объема требует усилия для того, чтобы раскрыть спавшиеся стенки альвеол в силу значительного поверхностного натяжения жидкости, покрывающей как их поверхность, так и поверхность дыхательных путей. Поэтому кривые, полученные при раздувании и спадении легких, не совпадают друг с другом, и их нелинейное изменение называется гистерезисом.
Легкие при спокойном дыхании никогда не спадаются полностью, поэтому кривая спадения описывает изменения легочного объема при изменении величины внутриплеврального давления в диапазоне от —2 см. водн. ст. до —10 см водн. ст. В норме легкие человека имеют высокую растяжимость (200 мл/см водн. ст.). Эластичность легочной ткани обусловлена свойствами соединительных волокон легочной ткани. С возрастом эти волокна, как правило, снижают тонус, что сопровождается увеличением растяжимости и уменьшением эластической тяги легких. При повреждении легочной ткани или при избыточном развитии в ней соединительной ткани (фиброз) легкие становятся плохо растяжимыми, величина их растяжимости снижается, что затрудняет выполнение вдоха и требует значительно большего усилия дыхательных мышц, чем в норме.
Легочная растяжимость обусловлена не только эластичностью легочной ткани, но и поверхностным натяжением слоя жидкости, покрывающей альвеолы. По сравнению с эластической тягой легких, влияние на величину легочной растяжимости во время дыхания фактора поверхностного натяжения слоя жидкости, покрывающей альвеолы, имеет более сложную природу.
Тонкий слой жидкости покрывает поверхность альвеол легких. Переходная граница между воздушной средой и жидкостью имеет поверхностное натяжение, которое формируется межмолекулярными силами и которое будет уменьшать площадь покрываемой молекулами поверхности. Однако миллионы альвеол легких, покрытых мономолекулярным слоем жидкости, не спадаются, поскольку эта жидкость содержит субстанции, которые в целом называются сурфактантом (поверхностно активный агент). Поверхностно активные агенты обладают свойством снижать поверхностное натяжение слоя жидкости в альвеолах легких на границе фаз воздух—жидкость, благодаря которому легкие становятся легко растяжимыми.
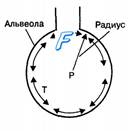
Рис. 10.7. Приложение закона Лапласа к изменению поверхностного натяжения слоя жидкости, покрывающего поверхность альвеол. Изменение радиуса альвеол изменяет в прямой зависимости величину поверхностного натяжения в альвеолах (Т). Давление (Р) внутри альвеол также варьирует при изменении их радиуса: уменьшается при вдохе и увеличивается при выдохе.
Альвеолярный эпителий состоит из плотно контактирующих между собой альвеолоцитов (пневмоцитов) I и II типа и покрыт мономолекулярным слоем сурфактанта, состоящего из фосфолипидов, белков и полисахаридов (глицерофосфолипиды 80 %, глицерол 10 %, белки 10 %). Синтез сурфактанта осуществляется альвеолоцитами II типа из компонентов плазмы крови. Основным компонентом сурфактанта является дипальмитоилфосфатидилхолин (более 50 % фосфолипидов сурфактанта), который адсорбируется на границе фаз жидкость—воздух с помощью белков сурфактанта SP-B и SP-C. Эти белки и глицерофосфолипиды уменьшают поверхностное натяжение слоя жидкости в миллионах альвеол и обеспечивают легочной ткани свойство высокой растяжимости. Поверхностное натяжение слоя жидкости, покрывающей альвеолы, изменяется в прямой зависимости от их радиуса (рис. 10.7). В легких сурфактант изменяет степень поверхностного натяжения поверхностного слоя жидкости в альвеолах при изменении их площади. Это обусловлено тем, что во время дыхательных движений количество сурфактанта в альвеолах остается постоянным. Поэтому при растяжении альвеол во время вдоха слой сурфактанта становится тоньше, что вызывает снижение его действия на поверхностное натяжение в альвеолах. При уменьшении объема альвеол во время выдоха молекулы сурфактанта начинают более плотно прилегать друг к другу и, увеличивая поверхностное давление, снижают поверхностное натяжение на границе фаз воздух—жидкость. Это препятствует спадению (коллапсу) альвеол во время экспирации, независимо от ее глубины. Сурфактант легких влияет на поверхностное натяжение слоя жидкости в альвеолах в зависимости не только от ее площади, но и от направления, в котором происходит изменение площади поверхностного слоя жидкости в альвеолах. Этот эффект сурфактанта называется гистерезисом (рис. 10.8).
Физиологический смысл эффекта заключается в следующем. При вдохе по мере увеличения объема легких под влиянием сурфактанта увеличивается натяжение поверхностного слоя жидкости в альвеолах, что препятствует растяжению легочной ткани и ограничивает глубину инспирации. Напротив, при выдохе поверхностное натяжение жидкости в альвеолах под влиянием сурфактанта уменьшается, но не исчезает полностью. Поэтому даже при самом глубоком выдохе в легких не происходит спадения, т. е. коллапса альвеол.
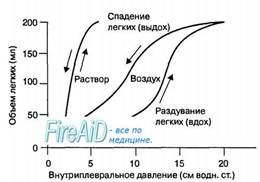
Рис. 10.8. Эффект поверхностного натяжения слоя жидкости на изменение объема легких в зависимости от внутриплеврального давления при раздувании легких солевым раствором и воздухом. Когда объем легких увеличивается за счет их наполнения солевым раствором, то в них отсутствуют поверхностное натяжение и феномен гистерезиса. Относительно интактных легких — площадь петли гистерезиса свидетельствует об увеличении поверхностного натяжения слоя жидкости в альвеолах при вдохе и снижении этой величины при выдохе.
В составе сурфактанта имеются белки типа SP-A и SP-D, благодаря которым сурфактант участвуют в местных иммунных реакциях, опосредуя фагоцитоз, поскольку на мембранах альвеолоцитов II типа и макрофагов имеются рецепторы SP-A. Бактериостатическая активность сурфактанта проявляется в том, что это вещество опсонизирует бактерии, которые затем легче фагоцитируются альвеолярными макрофагами. Кроме того, сурфактант активирует макрофаги и влияет на скорость их миграции в альвеолы из межальвеолярных перегородок. Сурфактант выполняет защитную роль в легких, предотвращая непосредственный контакт альвеолярного эпителия с частицами пыли, агентами инфекционного начала, которые достигают альвеол с вдыхаемым воздухом. Сурфактант способен обволакивать инородные частицы, которые затем транспортируются из респираторной зоны легкого в крупные дыхательные пути и удаляются из них со слизью. Наконец, сурфактант снижает поверхностное натяжение в альвеолах до близких к нулевым величинам и тем самым создает возможность расправления легких при первом вдохе новорожденного.
Растяжимость легких количественно характеризует растяжимость легочной ткани в любой момент изменения их объема в течение фазы вдоха и выдоха. Поэтому растяжимость представляет собой статическую характеристику эластических свойств легочной ткани. Однако во время дыхания возникает сопротивление движению аппарата внешнего дыхания, обусловливающее его динамические характеристики, среди которых наибольшее значение имеет сопротивление потоку воздуха при его движении через дыхательные пути легких.
На движение воздуха из внешней среды через дыхательные пути к альвеолам и в обратном направлении оказывает влияние градиент давления: при этом воздух движется из области высокого давления в область низкого давления. При вдохе давление воздуха в альвеолярном пространстве меньше, чем атмосферное, а при выдохе — наоборот. Сопротивление дыхательных путей потоку воздуха зависит от градиента давления между полостью рта и альвеолярным пространством.
Поток воздуха через дыхательные пути может быть ламинарным, турбулентным и переходным между этими типами. Воздух движется в дыхательных путях, в основном, ламинарным потоком, скорость которого выше в центре этих трубок и меньше вблизи их стенок. При ламинарном потоке воздуха его скорость линейно зависит от градиента давления вдоль дыхательных путей. В местах деления дыхательных путей (бифуркации) ламинарный поток воздуха переходит в турбулентный. При возникновении турбулентного потока в дыхательных путях возникает дыхательный шум, который может выслушиваться в легких с помощью стетоскопа. Сопротивление ламинарному потоку газа в трубе обусловлено ее диаметром. Поэтому, согласно закону Пуа-зейля величина сопротивления дыхательных путей потоку воздуха пропорциональна их диаметру, возведенному в четвертую степень. Поскольку сопротивление дыхательных путей находится в обратной зависимости от их диаметра в четвертой степени, то этот показатель самым существенным образом зависит от изменений диаметра воздухоносных путей, вызванных, например, выделением в них слизи из слизистой оболочки или сужением просвета бронхов. Общий диаметр сечения дыхательных путей возрастает в направлении от трахеи к периферии легкого и становится максимально большим в терминальных дыхательных путях, что вызывает резкое снижение сопротивления потоку воздуха и его скорости в этих отделах легких. Так, линейная скорость потока вдыхаемого воздуха в трахее и главных бронхах равна примерно 100 см/с. На границе воздухопроводящей и переходной зон дыхательных путей линейная скорость воздушного потока составляет около 1 см/с, в дыхательных бронхах она снижается до 0,2 см/с, а в альвеолярных ходах и мешочках — до 0,02 см/с. Столь низкая скорость воздушного потока в альвеолярных ходах и мешочках обусловливает в них незначительное сопротивление движущемуся воздуху и не сопровождается значимыми затратами энергии мышечного сокращения.
Напротив, наибольшее сопротивление дыхательных путей потоку воздуха возникает на уровне сегментарных бронхов в связи с наличием в их слизистой оболочке секреторного эпителия и хорошо развитого гладкомышечного слоя, т. е. факторов, которые в наибольшей степени влияют как на диаметр воздухоносных путей, так и на сопротивление в них потоку воздуха. В преодолении этого сопротивления заключается одна из функций дыхательных мышц.
В легких большинство дыхательных путей представляют собой эластичные трубки, за исключением трахеи и бронхов, стенки которых «укреплены» хрящевой тканью. Бронхиолы имеют высокоэластичные стенки, и диаметр их просвета может изменяться пассивно во время дыхательных движений. В обычных физиологических условиях при вдохе (как спокойном, так и глубоком) растяжение легочной ткани вызывает растяжение стенки мелких дыхательных путей. Согласно закону Пуазейля, незначительное увеличение радиуса дыхательных путей резко снижает в них сопротивление потоку воздуха. Поэтому при вдохе сопротивление дыхательных путей потоку воздуха не оказывает существенного влияния на силу сокращения дыхательных мышц. Напротив, при выдохе, особенно при глубоком и усиленном (форсированном) выдохе, диаметр мелких дыхательных путей уменьшается, что вызывает значительное увеличение сопротивления потоку воздуха в них. Влияние объема легких при выдохе на поток воздуха в дыхательных путях количественно характеризуется зависимостью «поток—объем». В клинической физиологии дыхания оценка этой зависимости является основным критерием типа и степени нарушения функции дыхательных путей.
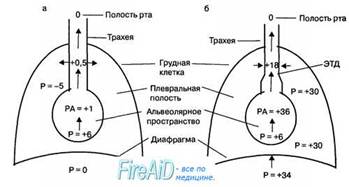
Рис. 10.9. Давление в дыхательных путях при выдохе. Вертикальными стрелками показаны величины давления, возникающие в дыхательных путях под влиянием комплайенса легких и грудной клетки. Горизонтальными стрелками в области дыхательных путей показано, что давление, оказываемое на стенки дыхательных путей, может увеличивать их просвет при спокойном выдохе (а) либо уменьшать их диаметр при глубоком выдохе (б) в том участке общей площади поперечного сечения мелких дыхательных путей, где сравниваются величины внутриплеврального и альвол и давления в дыхательных путях (эквипотенциальная точка — ЭПТ). Р —давление (см водн. ст.), РА — давление в альвеолах.
Зависимость «поток—объем» следующим образом характеризует влияние большого объема воздуха в легких на экспираторный поток воздуха в дыхательных путях (рис. 10.9). В момент, предшествующий началу выдоха, после глубокой инспирации в дыхательных путях отсутствует поток воздуха, а внутриплевральное давление равно —10 см водн. ст. С началом форсированной экспирации внутриплевральное давление возрастает примерно до +30 см водн. ст. относительно атмосферного давления, вызывая уменьшение радиуса как альвеол, так и мелких дыхательных путей. В этих условиях давление газов внутри альвеол становится выше, чем в плевральной полости, благодаря действию на стенки альвеол эластической тяги легких. В результате поток воздуха выходит из альвеолярного пространства по дыхательным путям во внешнюю среду по градиенту давления, который постепенно уменьшается в дыхательных путях по мере приближения к трахее. Спадению эластичных стенок бронхиол препятствует градиент давления воздуха между дыхательными путями и внутриплевральным давлением. Однако в некоторой точке дыхательных путей (как правило, в области бронхиол) этот градиент давления становится равным нулю (эквипотенциальная точка давления) и стенки дыхательных путей могут частично или полностью спадаться. В этих условиях продвижение воздуха по дыхательным путям может обеспечиваться только за счет увеличения силы сокращения (работы) внутренних межреберных мышц и мышц живота.
Снижение эластической тяги легких, например при эмфиземе легких, вызывает смещение ближе к альвеолярному пространству эквипотенциальной точки давления в дыхательных путях при выдохе, и, таким образом, блокируется выход воздуха непосредственно из альвеол. Дыхательные шумы, которые возникают в легких у больных, обусловлены прохождением воздуха через спавшиеся мелкие дыхательные пути. Увеличение экспираторного усилия у таких пациентов повышает риск спадения мелких дыхательных путей и еще больше затрудняет выдох. При бронхиальной астме у пациентов дыхательные пути уменьшают свой просвет в результате сокращения гладких мышц стенки бронхиол. В этом случае увеличение сопротивления потоку воздуха в мелких дыхательных путях вызывает рост градиента давления вдоль дыхательных путей при вдохе и смещает эквипотенциальную точку ближе к альвеолярному пространству, вызывая коллапс дыхательных путей при выдохе. Усиление сокращения экспираторных мышц в фазу выдоха еще больше затрудняет выдох у пациентов вследствие уменьшения просвета мелких дыхательных путей.
Сокращение дыхательных мышц создает градиент давления по ходу дыхательных путей. При этом преодолевается эластическое сопротивление легких и грудной клетки, а также сопротивление дыхательных путей потоку воздуха. Наряду с этим последние два показателя позволяют измерять работу дыхательных мышц во время дыхательного цикла. Если принять, что величина работы (W) представляет собой произведение силы (F) на путь (х), то получим: W = F х х В дыхательной системе, в которой измеряемыми величинами являются дыхательный объем и внутриплевральное давление, сила сокращения дыхательных мышц приравнивается к развиваемому ими давлению (Р), которое они оказывают на площадь (А). Поэтому, подставляя выражение F = Р х А в формулу работы дыхательных мышц в течение дыхательного цикла, получим: W = Р х А хх. Поскольку величина А, умноженная на путь (х), в дыхательной системе представляет собой аналог дыхательного объема (V), то общая формула работы дыхательных мышц имеет вид: W = Р х V.
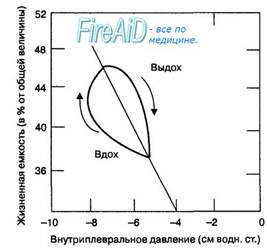
Рис. 10.10. Работа дыхательных мышц при спокойном дыхании. Изменения дыхательного объема (вертикальная ось) при вдохе и выдохе сопровождаются изменениями внутри-плеврального давления. При одновременной регистрации этих величин во время дыхательного цикла общая площадь петель дыхательный объем — внутриплевральное давление отражает количественно работу дыхательных мышц. Работа дыхательных мышц при вдохе больше, поскольку она затрачивается на преодоление эластического сопротивления легких. При выдохе работа дыхания минимальная, поскольку совершается за счет энергии эластической тяги легких, т. е. пассивно. Стрелками показаны изменения внутриплев-рального давления в течение фаз дыхательного цикла. Чем больше площадь петли, тем больше работа дыхательных мышц.
Работа дыхательных мышц при спокойном дыхании. При спокойном дыхании объем вдоха достигает максимум 1 л, а инспираторные мышцы совершают минимальную работу (рис. 10.10). Сокращение инспираторных мышц обеспечивает вдох, а выдох осуществляется пассивно за счет эластической тяги легких. В этих условиях сопротивление дыхательных путей при вдохе и выдохе не оказывает лимитирующего влияния на процесс внешнего дыхания. По мере увеличения глубины дыхания дыхательный объем формируется за счет объема функциональной остаточной емкости и резервного объема вдоха, а работа дыхания совершается против существенного нарастания поверхностного натяжения жидкости на поверхности альвеол. Поэтому чем глубже инспирация, тем большую работу совершают инспираторные мышцы. Во время выдоха, когда глубина дыхательных движений осуществляется в пределах объема жизненной емкости легких, объем легких возвращается пассивно к уровню функциональной остаточной емкости за счет эластической тяги легких, а в пределе функциональной остаточной емкости выдох происходит активно в результате сокращения мышц живота, которые при этом совершают работу.
Работа дыхательных мышц при глубоком дыхании. При глубоком дыхании на силу сокращения дыхательных мышц начинает оказывать влияние изменение диаметра дыхательных путей. Глубокий вдох вызывает расширение дыхательных путей и снижение сопротивления в них потоку вдыхаемого воздуха, поэтому работа инспираторных мышц обусловлена только величинами комплайенса легких и тканей грудной клетки. При глубоком выдохе, при котором в вьщыхаемом воздухе оказывается объем воздуха функциональной остаточной емкости, возникает сдавление мелких дыхательных путей высоким градиентом давления между дыхательными путями и внутриплевральным давлением. Существенное увеличение потока газов через дыхательные пути приводит к росту их сопротивления потоку воздуха, которое становится основным фактором, обусловливающим величину работы дыхания. Однако при глубоком дыхании механизмы регуляции диаметра дыхательных путей при участии вегетативной нервной системы способны минимизировать величину работы, которые выполняют дыхательные мышцы. Так, при глубоком дыхании за счет регулирующих влияний вегетативной нервной системы на гладкие мышцы дыхательных путей увеличивается их диаметр. В результате на сокращение дыхательных мышц затрачивается минимальное количество энергии. Например, при астме дыхание у пациентов становится медленным и глубоким, что снижает затраты энергии на преодоление сопротивления дыхательных путей потоку воздуха и уменьшает работу дыхательных мышц.
C 13.10








