 2018-02-13
2018-02-13 8223
8223БУТЕРБРОДНАЯ МОДЕЛЬ (белки – липиды – белки)
В 1935г. английские ученые Даниэли и Даусон высказали идею о послойном расположении в мембране молекул белков (темные слои в электронном микроскопе), которые залегают снаружи, и молекул липидов (светлый слой) – внутри. Длительное время существовало представление о едином трехслойном строении всех биологических мембран.
При детальном изучении мембраны с помощью электронного микроскопа оказалось, что светлый слой на самом деле представлен двумя слоями фосфолипидов – это билипидный слой, причем водорастворимые его участки – гидрофильные головки направлены к белковому слою, а нерастворимые (остатки жирных кислот) – гидрофобные хвосты обращены друг к другу.

Однако уже с середины 60-х годов начали накапливаться факты против унитарной «бутербродной» модели. В частности, по одним данным, не все мембраны имели четкую трехслойную структуру при электронно-микроскопическом исследовании; по другим – значительная часть мембранных белков имела глобулярную структуру, а не ламеллярную, как в постулируемой модели. Наконец, среди многочисленных моделей мембран, предложенных в середине 60-х годов, начали выделяться те, в которых доказывалось наличие гидрофобно-гидрофильных взаимодействий не только между липидными молекулами, но и между липидами и белками.
ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
В 1972г. Сингер и Николсон описали модель мембраны, которая получила широкое признание. Согласно этой модели молекулы белков не образуют сплошного слоя, а погружены в биполярный липидный слой на разную глубину в виде мозаики. Глобулы белковых молекул, подобно айсбергам, погружены в «океан»
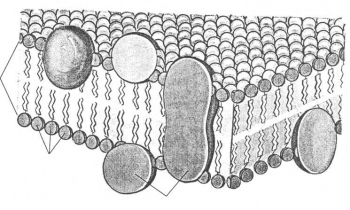
липидов: одни находятся на поверхности билипидного слоя – периферические белки, другие погружаются в него наполовину – полуинтегральные белки, третьи – интегральные белки – пронизывают его насквозь, формируя гидрофильные поры. Периферические белки, находясь на поверхности билипидного слоя, связаны с головками липидных молекул электростатическими взаимодействиями. Но они никогда не образуют сплошного слоя и, по сути дела, не являются белками собственно мембраны, а, скорее, связывают ее с надмембранной или субмембранной системой поверхностного аппарата клетки.
Основную роль в организации собственно мембраны играют интегральные и полуинтегральные (трансмембранные) белки, имеющие глобулярную структуру и связанные с липидной фазой гидрофильно-гидрофобными взаимодействиями. Молекулы белков, как и липиды, обладают амфипатричностью и своими гидрофобными участками взаимодействуют с гидрофобными хвостами билипидного слоя, а гидрофильные участки обращены к водной среде и образуют с водой водородные связи.
БЕЛКОВО-КРИСТАЛЛИЧЕСКАЯ МОДЕЛЬ (модель липопротеинового коврика)
Мембраны образованы переплетением липидных и белковых молекул, объединяющихся между собой на основе гидрофильно-
гидрофобных взаимодействий.

Белковые молекулы, как штифты, пронизывают слой липидов и выполняют в составе мембраны функцию каркаса. После обработки мембраны жирорастворимыми веществами белковый каркас сохраняется, что доказывает взаимосвязь между молекулами белков в мембране. По-видимому, эта модель реализуется лишь в отдельных специальных участках некоторых мембран, где требуется жесткая структура и тесные стабильные взаимоотношения между липидами и белками (например, в области расположения фермента Na-К –АТФ-азы).
Самой универсальной моделью, отвечающей термодинамическим принципам (принципам гидрофильно-гидрофобных взаимодействий), морфо-биохимическим и экспериментально-цитологическим данным, является жидкостно-мозаичная модель. Однако все три модели мембран не исключают друг друга и могут встречаться в разных участках одной и той же мембраны в зависимости от функциональных особенностей данного участка.
Функции мембран.
Функции
- барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемостьозначает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии по градиенту концентрации путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивает в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+). - матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
- механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней. - ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса. - маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.








