 2020-04-12
2020-04-12 805
805Все типы липопротеинов имеют сходное строение: гидрофобное ядро и гидрофильный слой на поверхности, образованный белками- апопротеинами и амфифильными молекулами липидов-фосфолипидами и холестеролом.
Хиломикроны.
Жиры, образовавшиеся в результате ресинтеза в клетках слизистой оболочки кишечника, упаковываются в ХМ. Основной апопротеин- белок апоВ-48. В аппарате Гольджи происходит формирование «незрелых» ХМ. По механизму экзоцитоза они выделяются в хилус, образующийся в лимфатической системе кишечных ворсинок, и через грудной лимфатический проток попадают в кровь. В лимфе и крови с ЛПВП на ХМ переносятся апопротеины Е и С- II; ХМ превращаются в «зрелые». ХМ транспортируют жиры к различным тканям, где он утилизируется, поэтому концентрация ХМ в крови постепенно снижается.
ХМ на 85% состоят из триацилглицеридов.
ЛПОНП
Синтезируются в печени и секретируются в кровь.
Состав: жиры, холестерол, фосфолипиды и белок апоВ-100. В крови на ЛПОНП действует ЛП-липаза. Жирные кислоты поступают в ткани, в частности в адипоциты.В процессе удаления жиров из ЛПОНП ЛПОНП сначала превращается ЛППП, а затем в ЛПНП.
ЛПОНП на 55% состоят из триацилглицеридов.
ЛПНП
Образуются в кровииз ЛПОНП и ЛППП.
ЛПНП на 7% состоят из триацилглицеридов.
Функция: транспорт холестерола в ткани
Апопротеин-В100
ЛПВП
ЛПВП на 3% состоят из триацилглицеридов.
Функция: удаление избытка холестерола из клеток и других ЛП. Донор апопротеинов А, С-II
Образование в клетках печени
Апопротеины А, С-II, E.
Окисление насыщенных жирных кислот с четным числом углеродных атомов: активация в цитоплазме, транспорт в митохондрии, b- окисление, связь с ЛКЦ и ЦПЭ (на примере окисления пальмитата), биоэнергетика процесса.
1. активирование происходит на поверхности митохондрий
CH3(CH2)14COOH 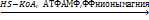 CH3-(CH2)14-CO-S-KoA
CH3-(CH2)14-CO-S-KoA
Фермент: Ацил-КоА-синтаза
2. транспорт через митохондриальную мембрану (карнитин)
CH3-(CH2)14-CO-S-KoA + (СH3)3-N-CH2-CH(OH) –CH2-COOH 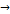
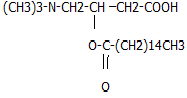 + HS-KoA
+ HS-KoA
ацилкарнитин
Фермент: карнитинацил-трансфераза
3. первоедегидрирование
Ацил-КоА подвергается фермент дегидрированию с образованием Еноил-КоА(с двойной связью между С2 И С3).
CH3-(CH2)14-CH2-CH2-CO-S-KoA + ФАД→ФАДН2 +CH3-(CH2)14-CH=CH-CO-S-KoA
фермент: ацил-КоА-дегидрогеназа
4. гидратация (из Еноила образуется 3-гидроксиацил-КоА)
CH3-(CH2)14-CH=CH-CO-S-KoA + H2O→ CH3-(CH2)14-CH(OH)-CH2-CO-S-KoA
Фермент: еноил-КоА-гидратаза
5. второе дегидрирование (образуется Оксоацил-КоА)
CH3-(CH2)14-CH(OH)-CH2-CO-S-KoA+ НАД+ → НАДН+Н+ + CH3-(CH2)14-CO-CH2-CO-S-KoA
Фермент: 3-гидроксиацил-КоА-дегидрогеназа
7. тиолазная реакция(В результате образуется ацил-КоА, укороченный на 2 углеродных атома и ацетил-КоА (в цикл трикарбоновых кислот)
CH3-(CH2)14-CO-CH2-CO-S-KoA→СH3-CO-S-KoA+ CH3-(CH)12-CH2-CH2-CO-S-KoA
Фермент: ацетил-КоА-ацетиттрансфераза
Биоэнергетика:
n= 7*16-6= 106 АТФ








