 2020-05-13
2020-05-13 96
96Https://zinref.ru/000_uchebniki/00500biologia/049_mutaciya-v-ontogene-destabilizaciya-genoma-ovanie-2005/000.htm
Мутация в онтогене – дестабилизация генома – формообразование (Б.Ф.Чадов, Е.В.Чадова, Е.А.Хоцкина, Н.Б.Федорова)
УДК: 575
Мутация в онтогене – дестабилизация генома – формообразование*
Б.Ф.Чадов, Е.В.Чадова, Е.А.Хоцкина, Н.Б.Федорова
Институт цитологии и генетики СО РАН, г. Новосибирск,
факс (3832)331278, e-mail: chadov@bionet.nsc.ru
Аннотация
Описывается феноменология дестабилизации генома у мутантов Drosophila melanogaster, содержащих мутации онтогенов (регуляторных генов, управляющих онтогенезом). Дестабилизация выражается в: 1) потере или снижении летальности ранее летальных мутаций; 2) потере проявления доминантных мутаций в оппозитном гомологе; 3) хромосомной нестабильности в виде потери Х-хромосомы в половых и соматических клетках; 4) образовании новых мутаций (с полной и неполной пенетрантностью), а также т.н. диморфных мутаций; 5) появлении единичных и массовых модификаций; 6) серийном характере появления мутаций; 7) массовом образовании морфозов. Предполагается, что причиной перехода генома из стационарного состояния в нестационарное является появление мутации в онтогене. К образованию нового вида может привести формирование новации – генетически обусловленного комплексного изменения фенотипа в период пребывания генома в нестационарном состоянии.
Таблиц – 4, рисунков – 7.
Введение
В 30-х годах прошлого века Четвериков, Фишер и Райт соединили дарвиновскую эволюционную идею с генетикой [1, 2, 3]. Возникли т. н. «синтетическая» теория эволюции и популяционная генетика, как полигон для ее проверки. Процесс эволюции был представлен как «естественный отбор» генных вариантов (аллелей). В рамках стремительно развившейся популяционной генетики был накоплен обширный материал о генетическом полиморфизме и его динамике [4, 5]. Оказалось, однако, что изменения частот аллелей в популяциях носят обратимый характер. Не найдено финальной направленности процесса, не говоря уже о фактах совершившегося видообразования. Популяционные генетики склонны считать, что аллельное разнообразие, наблюдаемое в популяциях, служит, скорее, адаптации вида в пределах ареала, чем является материалом для биологической эволюции [6, 7, 8]. Вновь стали актуальными гипотезы номогенеза [9] и сальтационного видообразования [10, 11, 12, 13].
Выход из кризиса, в котором находится генетическая интерпретация эволюции, видится в том, чтобы в качестве материала для эволюции рассматривать не традиционные менделевские гены, обеспечивающие внутривидовую изменчивость, а гены «инвариантной» части генома [6, 7], обеспечивающие внутривидовое сходство [14, 15]. Авторами статьи с использованием средств прямой генетики разработаны методы получения мутаций по таким генам у дрозофилы [14, 16]. Первые исследования мутаций показали, что они принадлежат регуляторным сигнальным генам, осуществляющим процесс индивидуального развития [17, 18, 19]. Эти гены назвали онтогенами [15].
Мутации онтогенов обладают особенностями, делающие их пригодными для продолжительной и скрытой перестройки генетической системы. Именно такая, долгое время не подконтрольная отбору, перестройка и должна, по нашему мнению, обеспечивать видообразование. Одна из важнейших особенностей полученных мутаций - способность дестабилизировать геном.
Материалы и методы
Мутации в Х-хромосоме и аутосоме 2 получили в 2000 - 2001 годах [14, 16], в течение пятилетнего срока их поддерживали в виде лабораторных культур и наблюдали за ними. Формально – генетически эти мутации - условные доминантные летали. Показатель летального действия мутации в Х-хромосоме - отсутствие дочерей в скрещивании мутантного самца с самками линии «yellow». Показатель летального действия мутации в аутосоме 2 – отсутствие мутантного потомства обоего пола в таком же скрещивании. Мутации в Х-хромосоме (22 штуки) поддерживали двумя способами: в культуре со сцепленными Х- хромосомами и в культуре, содержащей инверсию Muller-5 в Х-хромосоме (рис.1).
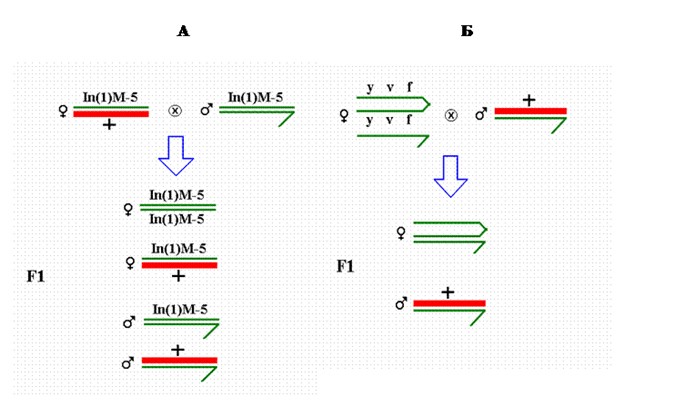
Рис. 1. Два способа поддержания мутаций онтогенов в Х-хромосоме: (А) в гетерозиготе у самок, содержащих инвертированную хромосому Muller-5 (In(1) M-5) и мутантную Х-хромосому (+, жирная линия) и (Б) в культуре со сцепленными Х- хромосомами (y v f/ y v f). В первом случае мутантную Х-хромосому получают дочери In(1)M-5/+ и сыновья +. Дочери In(1)M-5/ In(1)M-5 и сыновья In(1)M-5 мутантной Х-хромосомы не получают. Во втором случае – мутантную Х-хромосому получают сыновья, и не получают дочери.
Летальные мутации в аутосоме 2 (8 штук), поддерживали в гетерозиготе с инвертированной хромосомой In(2LR)Cy, Cy Bl L4 (рис.2). Эта хромосома содержала доминантные мутации Curly, Bristle и Lobe, которые отличаются устойчивым и отчетливо выраженным проявлением [20].
Каждую культуру поддерживали в 4-5 стаканчиках со стандартной средой для дрозофилы. Один - два раза в месяц её пересаживали на новую среду. При пересадке потомство просматривали под бинокулярной лупой. Новые фенотипические варианты, включая морфозы, документировали с помощью цифровой видеокамеры. В большинстве случаев от особи с необычным фенотипом пытались сделать отводок, скрещивая ее с сибсами. В исходной культуре, давшей новый фенотип, в следующем поколении целенаправленно искали повторного появления этого фенотипа.
Проверку летальности проводили путем скрещивания самцов из мутантных культур с самками yellow [16].
Результаты
Разлеталивание»
Потеря летального действия мутаций была обнаружена случайно в 2001 году в опытах по определению частоты доминантных леталей. Самцы из культур 1, 3, 5, 27 и 33, ведущихся на сцепленных Х-хромосомах, в скрещивании с самками yellow стали давать дочерей (табл.1). В последующие годы летальность проверили в более представительных выборках потомства. Пять вышеназванных мутаций, проверенные в 2002 и в 2004 годах, показали, что они действительно перестали быть леталями. В 2002 г. потеряли летальное действие еще 3 мутации (№№ 29, 38, 41), а в 2004 г. – ещё одна (№35). В некоторых культурах летальность не исчезла, но снизилась. В потомстве стали возникали дочери, хотя их доля не достигала положенного уровня 50% (№№ 7, 9, 10, 11). Всего из 22 летальных мутаций за период 2000-2004 гг. 9 мутаций полностью потеряли летальность, а 5 перешли в разряд полулеталей.
Таблица 1
Потеря летального проявления мутаций, полученных в 2000 году
(поддержание в культуре «attached –X»)
| Номер культу-ры | 2000 г. | 2001 г. | 2002 г. | 2004 г. | |||||
| Всего потом-ства | Доля дочерей | Всего потом-ства | Доля дочерей | Всего потом-ства | Доля дочерей | Всего потом-ства | Доля дочерей | ||
| 1 | 191 | 0.00 | 13 | *0.46 | 199 | *0.42 | 77 | *0.52 | |
| 2 | 435 | 0.00 | 4 | 0.00 | 259 | 0.02 | 36 | 0.03 | |
| 3 | 180 | 0.00 | 20 | *0.45 | 311 | *0.43 | 95 | *0.50 | |
| 5 | 303 | 0.02 | 33 | *0.45 | 265 | *0.60 | 83 | *0.41 | |
| 6 | 283 | 0.02 | 2 | 0.00 | 111 | 0.02 | 39 | 0.05 | |
| 7 | 100 | 0.00 | 3 | 0.00 | 44 | **0.27 | 63 | **0.40 | |
| 8 | 216 | 0.07 | 5 | 0.00 | 90 | 0.09 | 49 | **0.14 | |
| 9 | 529 | 0.00 | 7 | 0.00 | 169 | **0.21 | 81 | 0.04 | |
| 10 | 297 | 0.04 | 7 | 0.00 | 69 | **0.30 | 57 | **0.26 | |
| 11 | 409 | 0.06 | 4 | 0.00 | 82 | **0.18 | 55 | **0.16 | |
| 26 | 89 | 0.01 | - | 0.00 | 175 | 0.07 | 40 | 0.02 | |
| 27 | 161 | 0.00 | 29 | *0.69 | 113 | *0.56 | 92 | *0.49 | |
| 29 | 76 | 0.00 | 4 | 0.00 | 171 | *0.54 | 80 | *0.51 | |
| 30 | 115 | 0.00 | 8 | 0.00 | 109 | 0.02 | 71 | 0.00 | |
| 31 | 189 | 0.00 | 8 | 0.00 | 138 | 0.01 | 70 | 0.03 | |
| 32 | 198 | 0.00 | 4 | 0.00 | 74 | 0.00 | 53 | 0.02 | |
| 33 | 234 | 0.00 | 23 | *0.52 | 214 | *0.56 | 88 | *0.51 | |
| 34 | 198 | 0.00 | - | 0.00 | 62 | 0.00 | 54 | 0.02 | |
| 35 | 115 | 0.04 | 12 | 0.00 | 162 | **0.13 | 83 | *0.48 | |
| 36 | 110 | 0.01 | 5 | 0.00 | 106 | 0.02 | 54 | 0.07 | |
| 38 | 84 | 0.01 | 3 | 0.00 | 80 | *0.56 | 51 | **0.33 | |
| 41 | 100 | 0.01 | 5 | 0.00 | 331 | *0.49 | 106 | *0.52 | |
* - потеря летального действия мутации
** - снижение летального действия мутации
Процесс потери летальности мутаций №№ 1-41 происходил и в культурах, ведущихся в гетерозиготе с хромосомой Muller-5 (табл.2). За то же период времени полностью потеряли летальность 7 мутаций (№№ 3, 5, 9, 11, 32, 34, 38), снизили летальность – 8 (№№ 6, 7, 8, 10, 30, 31, 33, 36). Различие в способах поддержания мутаций определенно сказывалось на процессе разлеталивания. В культуре Muller-5 летали сохранялись более устойчиво. Разлеталивание мутаций №3, 5, 38 прошло при обоих способах поддержания, мутации № №2, 6, 30, 31 при обоих способах поддержания не утратили летальности. Для большинства мутаций (16 из 22) сохранение летальности и ее потеря не были скоординированы в культурах Muller-5 и «сцепленные Х-хромосомы»
В 2002 г., после того как была обнаружена потеря и резкое ослабление летальности 10 культур, поддерживающихся на сцепленных Х-хромосомах, их продублировали новыми культурами, полученными от самцов, взятых из соответствующих культур Muller-5.
К 2004 году 7 из них, теперь ведущихся на сцепленных Х-хромосомах, опять потеряли летальное действие (табл.3). Остались леталями три (№№ 27, 29, 41). Эти мутации остались леталями и при поддержании на Muller-5 (табл.2).
Таблица 2
Потеря летального действия мутаций, полученных в 2000 году и поддерживающихся в культурах «attached-X» и «Muller-5»
| Номер культуры самца | 2000 г. | 2004 г. attached-X | 2004 г. Muller-5 | |||
| Всего потомства | Доля дочерей | Всего потомства | Доля дочерей | Всего потомства | Доля дочерей | |
| 1 | 191 | 0.00 | 77 | 0.52 | 77 | 0.06 |
| 2 | 435 | 0.00 | 36 | 0.03 | 89 | 0.08 |
| 3 | 180 | 0.00 | 95 | *0.50 | 77 | *0.48 |
| 5 | 303 | 0.02 | 83 | *0.41 | 64 | *0.50 |
| 6 | 283 | 0.02 | 39 | 0.05 | 31 | **0.10 |
| 7 | 100 | 0.00 | 63 | **0.40 | 11 | **0.18 |
| 8 | 216 | 0.07 | 49 | **0.14 | 68 | **0.19 |
| 9 | 529 | 0.00 | 81 | 0.04 | 118 | *0.54 |
| 10 | 297 | 0.04 | 57 | **0.26 | 79 | **0.42 |
| 11 | 409 | 0.06 | 55 | **0.16 | 86 | *0.46 |
| 26 | 89 | 0.01 | 40 | 0.02 | - | - |
| 27 | 161 | 0.00 | 92 | *0.49 | 55 | 0.04 |
| 29 | 76 | 0.00 | 80 | *0.51 | 34 | 0.06 |
| 30 | 115 | 0.00 | 71 | 0.00 | 60 | **0.10 |
| 31 | 189 | 0.00 | 70 | 0.03 | 50 | **0.10 |
| 32 | 198 | 0.00 | 53 | 0.02 | 43 | *0.54 |
| 33 | 234 | 0.00 | 88 | *0.51 | 48 | **0.10 |
| 34 | 198 | 0.00 | 54 | 0.02 | 67 | *0.45 |
| 35 | 115 | 0.04 | 83 | *0.48 | 44 | **0.14 |
| 36 | 110 | 0.01 | 54 | 0.07 | 30 | **0.27 |
| 38 | 84 | 0.01 | 51 | **0.33 | 84 | *0.50 |
| 41 | 100 | 0.01 | 106 | *0.52 | 93 | 0.06 |
* - потеря летального действия мутации
** - снижение летального действия мутации
Полученные данные позволяют считать, что в культурах идет процесс изменения свойств мутаций. Он зависит как от самой мутации, так и от геномного окружения.
Таблица 3
Потеря летального проявления мутаций, содержавшихся первоначально в культуре Muller-5, а затем в культуре C(1)DX, y w f
| Номер культуры самца | Дочерей «+» | Сыновей yellow | Всего потомства | Доля дочерей |
| 1 | 53 | 65 | 118 | 0.45 |
| 3 | 44 | 83 | 127 | 0.35 |
| 5 | 62 | 49 | 111 | 0.56 |
| 7 | 72 | 60 | 132 | 0.54 |
| 11 | 33 | 34 | 67 | 0.49 |
| 27* | 0 | 58 | 58 | 0 |
| 29* | 3 | 66 | 69 | 0.04 |
| 33 | 47 | 61 | 108 | 0.44 |
| 38 | 66 | 60 | 126 | 0.52 |
| 41* | 4 | 47 | 51 | 0.08 |
* - мутации, не потерявшие летального действия
Потеря проявления доминантной мутации в оппозитной хромосоме
Летальные мутации в аутосоме 2, поддерживающиеся в гетерозиготе с инвертированной хромосомой In(2LR)Cy, Cy Bl L4, характеризуются «потерей проявления» доминантных мутаций Cy, Bl и L4 в инвертированной хромосоме (рис.2).








