 2020-05-13
2020-05-13 116
116В культурах мутаций появляются волны фенокопий. В одном или нескольких поколениях воспроизводится тот или иной фенотип известных мутаций: black, purple, brown, trident, abnormal abdomen, Notch, yellow, Dichaete и др. (рис. 5). В большинстве случаев появляется сразу несколько особей (от 3 до 10) нового фенотипа. В следующей генерации, как правило, число таких особей увеличивается. Однако через несколько поколений они начинают постепенно исчезать из потомства. Попытка закрепить новый фенотип в отводке оказывается безуспешной. Например, одной из часто возникающих фенокопий является наличие у мух меланомоподобных образований (рис. 5 а). Удавалось

Рис. 5. Модификации в культурах, содержащих мутацию в онтогене. а) «меланома» участок ткани темного цвета; б) «пузырчатое» крыло; в) «бледные карлики» - мухи малого размера и бледно-желтой окраски; г) вертикально загнутые крылья; д) «укороченные» крылья, крылья изменены и уменьшены вплоть до полного отсутствия; е) «карлик» - самец уменьшен в размере, нормально окрашен, сверху - самец нормального размера; ж) самка темной окраски, справа самец нормальной окраски; з) три особи с дефектом фасеточной части глаза.
получить отводки со 100% наличием «меланомы» у потомка, однако через некоторое количество поколений фенотип терялся окончательно.
В культурах мутаций периодически появляются мелкие особи. Вывести культуру с таким фенотипом, несмотря на неоднократные попытки, не удалось. Периодически возникают резкие нарушения в соотношении полов. Эти изменения ведут себя так же, как было указано выше для фенокопий.
6. Массовое образование морфозов .
Выщепление новых фенотипов в культурах мутаций онтогенов происходит на фоне образования разнообразных односторонних морфозов у потомков мутантов, о чем уже ранее сообщалось (15, 18, 23). По частоте встречаемости в культурах морфозы намного превосходят мутации и модификации.
О частоте образования эндогенных мутаций и модификаций у мутантов по онтогенам
Приведенные данные по образованию новых фенотипов (мутаций, модификаций и морфозов) основаны на наблюдениях за культурами мутаций при их поддержании или при участии в экспериментах. Строгих оценок частот нет, поскольку сами факты были неожиданными. Вместе с тем можно утверждать, что: 1) частоты обнаруженных событий на несколько порядков величин выше, чем в культурах обычных мутаций дрозофилы; 2) процесс образования новых фенотипов неравномерен, он наиболее интенсивен сразу после получения мутации и в случае ее гибридизации с иной культурой; 3) существует тенденция к затуханию образования новых форм в процессе поддержания мутанта в культуре.
Обсуждение
Наблюдение за культурами, содержащими мутации онтогенов, показало, что мутантные геномы находятся в состоянии нестабильности. Нестабильность проявляется в нескольких формах. Разлеталивание говорит о нестабильности самих мутировавших локусов. Непроявление доминантных мутаций в оппозитной хромосоме и образование новых мутаций указывают на распространение нестабильности на другие локусы. Потеря и нерасхождение Х-хромосом свидетельствуют о нарушении мейоза, а образование мозаиков и гинандроморфов – о нарушении митоза. О неблагополучии генетических процессов в митотически делящихся клетках говорит и образование морфозов. Морфозы представляют собой односторонние нарушения хода онтогенеза в ограниченной группе клеток развивающегося организма.
Нестабильность касается как структурных генов (образование фенотипически известных мутаций), так и регуляторных генов, управляющих онтогенезом (онтогенов). Образование сложных морфологических структур в неположенном месте (морфозов) объясняется запуском в не предназначенных для этого клетках цепи автоматически следующих друг за другом онтогенетических событий.
Факты образования фенотипически известных мутаций со стойким наследованием в чреде поколений позволяет считать, что нарушения могут выражаться в изменении первичной последовательности ДНК. Однако нарушения могут и не затрагивать первичную структуру. Осуществляется вариант мутации только на одно поколение (типичная фенокопия) или вариант «транзитной» мутации на несколько поколений (волны фенокопий). В последнем случае должно происходить временное изменение участка ДНК в генеративном пути, которое передается потомкам в некотором числе поколений, а затем исчезает.
Полученная картина дестабилизации напоминает нестабильность, вызываемую перемещением мобильных элементов[24]. Новыми моментами являются два: 1) пусковой момент нестабильности - мутация в онтогене и 2) нестабильность распространяется, прежде всего, на онтогены. Тот факт, что нестабильность провоцировалась каждой из полученных мутаций, говорит о том, что состояние нестабильности является биологически узаконенной формой состояния генома. Распространение нестабильности в первую очередь на онтогены позволяет предполагать, что состояние нестабильности предназначается для поиска вариантов переустройства генома.
Генетическая расшифровка онтогенеза делает первые шаги и вопрос о перепрограммировании онтогенеза в процессе филогенеза пока далек от решения. Полученные данные, однако, позволяют утверждать, что мутагенез по онтогенам создает реальную возможность для переустройства генома.
Структурные гены находятся на концах регуляторных цепей онтогенеза (рис.6), существующие в природе их мутационные варианты не нарушают общей композиции организма. Классическая генетика показывает, насколько широк диапазон мутационной изменчивости по этим генам. По причине расположения на концах регуляторных путей и по причине уникальности структурных генов, мутации структурных генов, как правило, изменяют фенотип. Они становятся достоянием отбора. Отбор оставляет тех из них, которые своим присутствием не нарушают ход онтогенеза. Это значит, что среди мутаций, имеющих фенотипическое проявление и прошедших отбор, не окажется ни одной, которая представляла бы из себя серьезное изменение онтогенеза и могла бы служить образованию нового вида.
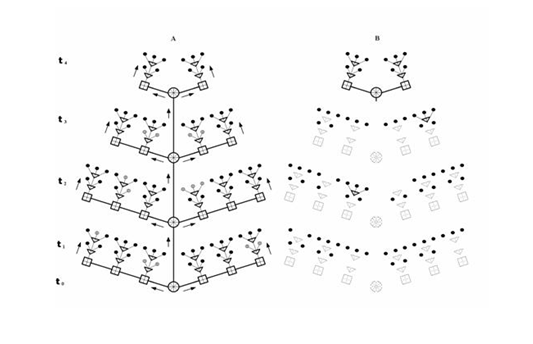
Рис. 6. Генетическая модель онтогенеза. А – гены и сигнальные пути. Геном особи состоит из структурных генов (кружки черного цвета) и онтогенов разных рангов (светлые круги, квадраты и треугольники). Онтоген представлен набором цис-аллелей (разделение значков на сектора). t0-4 – стадии онтогенеза. Активация генома идет по регламентированной системе сигнальных путей (линии между генами, стрелки). Сигнальные пути завершаются включением структурных генов. Онтогенез представляет собой процесс последовательного включения онтогенов разного ранга по принципу эстафеты. При переходе от предыдущей стадии онтогенеза к последующей стадии отключаются онтогены, работавшие на предыдущей стадии, а также структурные гены, обеспечивавшие появление презумптивных структур (заштрихованные кружки). Б – онтогенез на одной из последних стадий (t4). Пунктиром показаны отключенные онтогены и не действующие сигнальные пути. Остается включенной большая часть структурных генов и некоторые онтогены, близкие к ним по времени включения (онтогены стволовых клеток).
Мутации онтогенов возникают в организме по тем же законам, что и мутации структурных генов. Однако в отличие от последних они имеют более прямое отношение к механизму онтогенеза и к тому же могут находиться в геноме, не проявляясь фенотипически. Это происходит потому, что онтогены устроены по принципу кассеты цис-аллелей (рис.7). Мутация сохраняется в организме, у которого в активном состоянии находится нормальный цис-аллель, а мутантный цис-аллель - в не активном состоянии. Устройство онтогена в виде кассеты цис-аллелей позволяет иметь в геноме и работающую программу онтогенеза, и не работающую «теневую» программу, и части некой новой программы в стадии становления. В качестве иллюстрации возможности сосуществования в геноме нескольких программ развития приведем онтогенез у двуполых видов. В геноме каждого представителя вида генетически представлены и женская, и мужская программы развития, однако в активном состоянии находится одна.
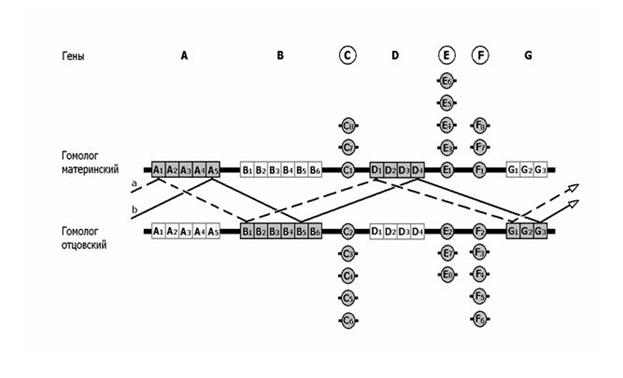
Рис 7. Схема генома диплоидного организма. C, E, F (в кружках) – уникальные структурные гены, в популяции существуют аллели этих генов (например, С1-С6); в диплоидном геноме не более двух аллеляей. У каждого представителя вида активны оба аллеля (заштрихованы). Онтогены A, B, D, G состоят из кассет цис-аллелей (например, А1-А5). У конкретного организма активен только один цис-аллель онтогена и только в одном из гомологов (заштрихованные кассеты). В зависимости от включения того или иного цис-аллеля реализуется конкретный вариант онтогенеза. a и b – варианты онтогенеза. Мутация в цис-аллеле D1 при нормальном развитии (вариант а) окажется летальной, но при развитии по варианту b ее действие не проявится.
Мутации онтогенов, обладая возможностью находиться в геноме без фенотипического проявления, могут накапливаться в нем. Будучи по отдельности летальными в активном состоянии, все вместе они могут утратить летальность. Это может произойти, если, соединяясь друг с другом, они создадут новую регуляторную цепь, вписывающуюся в ход онтогенеза. Не исключены и другие возможности реорганизации регуляторных цепей при наличии поврежденных звеньев. На уровне фенотипа новый регуляторный ансамбль вызовет появление «новации» (< innovation >) - нового морфологического или функционального дополнения, которое с помощью мутаций нельзя разложить на составные части. Тот факт, что мутация в онтогене запускает процесс нестабильности во всем геноме, указывает на то, что в живом организме существует путь достижения стабильного состояния способом активного, но скрытого до поры до времени переустройства генома.
Все мутанты по онтогенам, которые находились в опыте, существовали как живые организмы только потому, что мутантные аллели находились у них в неактивном "спящем" состоянии. Это означает, что есть способ, с помощью которого генетическая система обнаруживает присутствие не работающего мутантного аллеля, для того чтобы отреагировать на это дестабилизацией генома.
Предполагаем, что механизм дестабилизации состоит в инициации перемещения мобильных элементов в геноме. Полагаем, что система регуляторных генов, управляющих онтогенезом (онтогенов), является очень подвижной, а ее деятельность тесно связана с мобильными элементами. Перемещение последних по онтогенам является заурядным событием. Полученные данные позволяют считать, что перемещение мобильных элементов чаще всего происходит по ходу регуляторных цепей: от онтогена высшего порядка к низшему, далее - к структурным генам. Случаи повторного образования одних и тех же мутаций в культуре мутанта, образование мутантов пучками и одновременного образования мутантов в отдельно ведущихся семьях позволяет говорить о «рассылке мобильных элементов по адресам» («mailing» ), а не о хаотическом их перемещении, на что ранее указывали и другие авторы [25, 26].
Давно было замечено, что воздействие мутагенными факторами не только приводит к образованию мутаций в классическом генетическом смысле, но и вызывает «дестабилизацию» генома [27]. Она выявляется как у мутантов, так и не мутантов. У мутантов, к примеру, возрастает частота образования новых мутантных форм, появляются модификации, повышается вариабильность фенотипа [28, 29]. Нестабильность генома человека и животных после облучения привлекает внимание радиобиологов [30, 31]. Наши данные на дрозофиле, позволяют предполагать, что причиной нестабильности, вызванной внешними воздействиями, являются мутации в онтогенах, которые возникли при облучении и попали к потомкам.
Полученные данные являются еще одним аргументом в пользу представления о том, что главным биологическим предназначением мобильных элементов является участие в работе регуляторной части генома [32]. Их роль в эволюционном преобразовании генома [12, 33-35] становится все более определенной. Она сводится не столько повышению общей частоты выхода мутаций, сколько к переводу генома из стационарного состояния в не стационарное, в котором происходит специфическая, эндогенная по природе, перестройка регуляторной системы онтогенеза.
Работа выполнена при финансовой поддержке РФФИ: грант № 04-04-48100.
Литература
1. Четвериков С.С. О некоторых моментах эволюционного процесса с точки зрения современной генетики \\ Журн. эксперим. биологии. 1926. №1. С.3-54.
2. Fischer R.A. The general theory of natural selection. Oxford: Clarendon press, 1930. 272 p.
3. Wright S. Evolution in Mendelian populations // Genetics 1931.Vol. 16. P.97-159.
4. Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н..В. Очерк учения о популяции. М.: Наука, 1973, 277 с.
5. Dobzhansky Th., Ayala F.J., Stebbins G.L.. et al. Evolution. San Francisco: Freeman, 1977. 572 p.
6. Алтухов Ю.П., Рычков Ю.Г. Генетический мономофизм вида и его биологическое значение // Журн. общ. биологии. 1972. Т. 33. №3. С.281-300.
7. Алтухов Ю.П. Генетические процессы в популяциях. Москва, ИКЦ «Академкнига», 2003, 431с.
8. Carson A. The genetics of speciation at the diploid level. Amer. Nat, 1975, V.109, Р. 83 - 95.
9. Берг Л.С. Труды по теории эволюции. Л.: Наука, 1977.
10. Goldschmidt R.B. Evolution as viewed by one geneticist // Amer. Sci. 1952. Vol.40. P.84-98.
11. Стегний В.Н. Архитектоника генома, системные мутации и эволюция. Новосибирск: Изд-во Новосибирского ун-та, 1993, 110 с.
12. Евгеньев М.Б., Е.С. Зеленцова, Е.С. Полуэктова и др. Инвазия мобильных элементов – причина взрывного сальтационного видообразования. В кн.: Эволюционная биология. Материалы конференции «Проблема вида и видообразование»/ Под ред. В.Н. Стегния. Томск: Томский государственный университет. Т.1, 2001, 396 с.
13. Гродницкий Д.Л. Эпигенетическая теория эволюции как возможная основа нового эволюционного синтеза. Журн. общ. биологии, 2001, Т.62, N2, С.99-109.
14. Чадов Б.Ф. Мутации, способные инициировать видообразование. В кн. Эволюционная биология: Материалы конференции «Проблема вида и видообразование»/ Под ред. В.Н. Стегния. Томск: Томский государственный университет, Т.1, 2001. С. 138-162.
15. Чадов Б.Ф., Е.В. Чадова, С.А. Копыл, Е.В. Артемова, Е.А. Хоцкина, Н.Б. Фёдорова. От генетики внутривидовых отличий к генетике внутривидового сходства // Генетика. 2004. Т.40. № 9. С. 1157-1172.
16. Чадов Б.Ф., Чадова Е.В., Копыл С.А., Федорова Н.Б. Новый класс мутаций у Drosophila melanogaster // Доклады РАН. 2000. Т.373. №5. С. 714- 717.
17. Чадов Б.Ф. «Образ» регуляторного гена в опытах на дрозофиле // Генетика. 2002. Т.38. N 7. С.869-880.
18. Чадов Б.Ф. Факультативные доминантные летали: генетика, онтогенез и филогенез. В кн. Эволюционная биология: Материалы II конференции «Проблема вида и видообразование»/ Под ред. В.Н. Стегния. Томск: Томский государственный университет, Т.2, 2002. С.118-142.
19. Чадов Б.Ф., Н.Б. Федорова. Элементарное событие онтогенеза.Докл. РАН. 2003. Т. 389. No 3. С. 408 – 412.
20. Lindsley D. L., Grell E.H. Genetic Variation of Drosophila melanogaster. Carnegie Inst. Wash. Publ., 1968. № 627. 472 P.
21. Bridges C.B. Nondisjunction as proof of the chromosome theory of heredity. Genetics. V.1. P.1-52, 107-162.
22. Федорова Н.Б., Е.А Хоцкина, Е.Ю Митрёнина, Б.Ф. Чадов. Хромосомная перестройка и видообразование: объяснение связи между событиями. Вестник ТГУ (в печати).
23. Чадов Б.Ф., Чадова Е.В., Копыл С.А., Хоцкина Е.А., Федорова Н.Б. Гены, управляющие онтогенезом: морфозы, фенокопии, диморфы и другие видимые проявления мутантных генов // Генетика, 2004. Т.40. № 3. С.353-365.
24. Хесин Р.Б. Непостоянство генома. М.: Наука, 1984. 472 с.
25. Ратнер В. А., Забанов С.А., Колесникова О.В., Васильева Л.А. Анализ множественных транспозиций МГЭ Dm412 в геноме дрозофилы при помощи теплового шока // Генетика.1992.Т.28. С. 68-86.
26. Забанов С.А., Л.А. Васильева, В.А. Ратнер. Индукция транспозиций МГЭ Dm412 при помощи гамма - облучения в изогенной линии Drosophila melanogaster // Генетика. 1995. Т.31. С.798- 803.
27. Дубинин Н.П. Потенциальные изменения в ДНК и мутации (молекулярная цитогенетика). Москва: Наука. 1978. 244 с.
28. Сидорова К.К. Генетика мутантов гороха. Новосибирск, Наука. 1981. 168 с.
29. Сидорова К.К. Естественная и индуцированная мутабильность мутантов гороха. В сб. "«Современные концепции эволюционной генетики". ч. II, 1997. С. 319-321.
30. Kiefer J. Radiation biology: glory of the past – changes and challenges of the future. In: Современные проблемы радиобиологии, радиоэкологии и эволюции. Дубна: ОИЯИ, 2001, С.130-139.
31. Лавренчук Г.И., Серкиз Я.И., Дудченко Т.Н., Ряполова И.Ю. Радиогенные эффекты малых доз радиации у потомков облученных клеток. В сб.: Современные проблемы радиобиологии, радиоэкологии и эволюции. Дубна: ОИЯИ, 2001, С.221-232.
32. Ратнер В.А., Л.А.Васильева. Мобильные генетические элементы (МГЭ): “Эгоистическая ДНК” или функциональные компоненты генома? В сб. "«Современные концепции эволюционной генетики". ч. II, 1997. С. 289-291.
33. Гвоздев В.А., Кайданов Л.З. Геномная изменчивость, обусловленная транспозициями мобильных элементов, и приспособленность особей Drosophila melanogaster // Журн. общей биологии. 1986. Т.47. С.51.
34. Di Franco C., Galuppi D., Junackovic N. Genomic distribution of transposable elements among individuals of an inbred Drosophila line // Transposable elements and evolution/ Ed. McDonald J. Dordrecht: Kluwer Acad. Publ., 1993.
35. Biemont C., Arnault C., Heizmann A., Ronsseray S. Massive changes in genomic localization of P element in an inbred line of Drosophila melanogaster // Naturwissenschaften. 1990. B.77. S. 485-488.
////////////////////////////////////////








