 2020-05-25
2020-05-25 155
155В центральной части рисунка показана локализация интрафузальных рецепторов и сухожильных рецепторов Гольджи. Справа показано мышечное веретено с несколькими интрафузаль- ными волокнами внутри него: эфферентный аксон принадлежит у-мотонейрону спинного мозга; чувствительные окончания афферентного нейрона возбуждаются при растяжении центральной части интрафузальных волокон. Слева изображены рецепторы Гольджи, активация которых происходит пропорционально росту напряжения мышцы и натяжению сухожильных коллагеновых волокон, сдавливающих чувствительные окончания.
виде быстрого ответного сокращения мышцы, а при медленном растяжении — тонический рефлекс, направленный на сохранение неизменной длины мышцы при постоянном растяжении. Тонические рефлексы необходимы для поддержания мышечного тонуса, под которым понимают сопротивление мышц растяжению какой-либо внешней силой. Мышечный тонус сохраняет положение тела и обеспечивает сопротивление силе тяжести, растягивающей мышцы-разгибатели. Изменения тонуса мышц позволяют удерживать и перемещать груз, сохранять равновесие при отклонениях туловища вперед, назад или в сторону.
Служащие в качестве рецепторов растяжения интрафузальные мышечные волокна значительно тоньше и короче остальных, т. е. экстрафузалъ- ных, образующих основную массу мышцы и обеспечивающих ее сокращения (рис. 4.17). Мышечные веретена обоими концами прикреплены полоской соединительной ткани к пучку экстрафузальных волокон, поэтому растягиваются одновременно со всей мышцей. В средней части каждого интрафузального волокна содержатся окончания первичного афферентного нейрона, относящиеся к механорецепторам, которые реагируют на вызванную растяжением деформацию. Возбуждение сенсорных нейронов передается мотонейронам через моносинаптическую рефлекторную дугу, что приводит к рефлекторному сокращению растянутой мышцы.
При растяжении мышцы внешней силой происходит одновременная активация а-мотонейронов, иннервирующих основную массу мышечных волокон и у-мотонейронов, которые иннервируют только интрафузальные
 |

 |  |
волокна. Оба типа мотонейронов возбуждаются разными коллатералями одного и того же сенсорного нейрона, поэтому сокращения обоих типов мышечных волокон тоже происходит одновременно. Другие ветви афферентного аксона активируют тормозные интернейроны спинного мозга, которые реципрокно подавляют активность мотонейронов для мышц-антагонистов (рис. 4.18). Таким образом, в механизме рефлекса растяжения запрограммирована одновременная согласованная деятельность нескольких мышц, необходимая для совершения или предотвращения движения в определенном суставе.
Чувствительность интрафузальных волокон к растяжению изменяется при повышении или понижении активности у-мотонейронов. Чем чаще они возбуждаются, тем больше сократятся концевые участки интрафузальных волокон, в то время как их центральная часть, лишенная сократительных белков, будет вынуждена растягиваться. Небольшое ее растяжение существует в условиях физиологического покоя, что поддерживает возбуждение рецепторов мышечных веретен и тоническое сокращение мышц. Изменения активности у-мотонейронов, приводящие к изменениям тонуса
мышц, происходят под влиянием двигательных центров головного мозга. В зависимости от конкретной функциональной задачи избирательно активируются у-мотонейроны, иннервирующие разные группы мышц, например сгибатели или разгибатели. Соответственно этому тонус одних групп мышц будет повышаться, тогда как другие группы мышц останутся в прежнем состоянии или даже расслабятся, если произойдет торможение активности соответствующих у-мотонейронов.
4.3.2.3. Рефлекторная регуляция напряжения мышц
Сокращения мышцы сопряжены с изменениями не только ее длины, но и напряжения. Рефлекторная регуляция напряжения осуществляется с помощью сухожильных рецепторов Гольджи — немиелинизированных окончаний афферентного нейрона, которые расположены между коллагеновыми пучками сухожильных волокон в области их прикрепления к мышечным волокнам (см. рис. 4.17). Если интрафузальные волокна — датчики длины лежат параллельно экстрафузальным волокнам, составляющим основу мышцы, то сухожильные рецепторы соединены с ними последовательно. При нарастании напряжения в мышце коллагеновые волокна сухожилий натягиваются тем сильнее, чем большим становится напряжение. Пропорционально ему деформируются сухожильные рецепторы и возрастает частота потенциалов действия в афферентном нейроне. Окончания афферентных нейронов образуют синапсы с тормозными интернейронами, действующими на гомонимные (относящиеся к этой же мышце) мотонейроны (рис. 4.19). Такой механизм регуляции по принципу отрицательной обратной связи предупреждает неадекватное нарастание мышечного напряжения.

|
|
 Тормозные интернейроны, контролирующие возбуждение мотонейронов, а значит, и силу мышечных сокращений, получают афферентную информацию не только от сухожильных рецепторов Гольджи, но также от суставн ых ре це пторо в, высокочувствительных кожных рецепторов и нисходящих двигательных путей. Конвергенция кожных афферентов на тормозных интернейронах требуется в тех случаях, когда происходит ощупывание хрупких или мелких объектов и силу мышечных сокращений нужно согласовать с характером тактильных ощущений.
Тормозные интернейроны, контролирующие возбуждение мотонейронов, а значит, и силу мышечных сокращений, получают афферентную информацию не только от сухожильных рецепторов Гольджи, но также от суставн ых ре це пторо в, высокочувствительных кожных рецепторов и нисходящих двигательных путей. Конвергенция кожных афферентов на тормозных интернейронах требуется в тех случаях, когда происходит ощупывание хрупких или мелких объектов и силу мышечных сокращений нужно согласовать с характером тактильных ощущений.
 |

раздражителя, вы- сгибание ноги, на тонус мышц-разги-
бателей (+) и уменьшается тонус мышц-сгибателей (—), что позволяет опорной ноге удерживать массу тела. Тормозные интернейроны на схеме темные, возбуждающие — светлые.
Многие суставы способны производить движения в двух или даже трех плоскостях, а для этого требуются согласованные сокращения одних групп мышц при одновременном расслаблении других. Механизм сгибательного рефлекса включает взаимодействие возбуждающих и тормозных интернейронов спинного мозга с мотонейронами мышц-синергистов и антагонистов. Такое взаимодействие предусматривает решение трех задач: 1) непосредственный контроль каждой мышцы; 2) координация деятельности мышц, управляющих одним суставом; 3) координация деятельности мышц, необходимых для управления несколькими суставами.
Рефлекторное сгибание конечности часто сопровождается одновременным разгибанием контралатеральной конечности, которое тоже происходит рефлекторно (перекрестный разгибательный рефлекс). Так, например, человек, наступивший босой ногой на колючку, рефлекторно сгибает эту ногу, что приводит к росту нагрузки на опорную ногу. Немедленное рефлекторное повышение тонуса мышц-разгибателей на опорной ноге позволяет справиться с возросшей нагрузкой на нее. Осуще-
|
|
 |
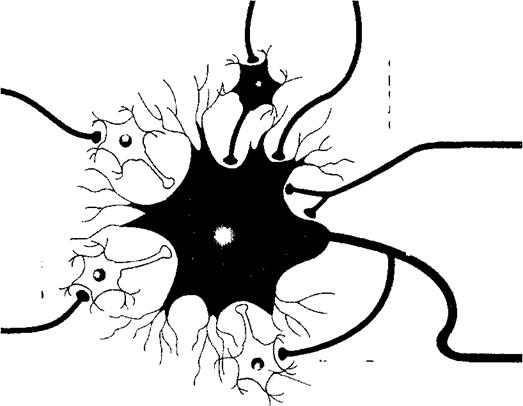
Мотонейрон могут возбуждать сенсорные нейроны, реагирующие на растяжение мышцы, возбуждающие интернейроны и нисходящие моторные пути головного мозга. На него могут действовать тормозные интернейроны, активированные рядом афферентных нейронов, а также клетки Реншоу. Схема показывает лишь малую часть реальных влияний на мотонейрон.
ствление перекрестного разгибательного рефлекса обеспечивается контралатеральной проекцией аксонов, берущих начало от интернейронов, которые возбуждаются при поступлении в спинной мозг сигналов от болевых рецепторов. В результате на противоположной стороне спинного мозга возбуждаются мотонейроны мышц-разгибателей и тормозятся мотонейроны сгибателей, т. е. наблюдается картина, прямо противоположная происходящему при сгибательном рефлексе.
Интернейроны, участвующие в образовании рефлекторных дуг сгибательных и разгибательных рефлексов, активируются окончаниями сенсорных нейронов разных модальностей. К ним конвергируют афференты от поверхности кожи, мышц, суставов, кроме того, они находятся под контролем нисходящих двигательных путей головного мозга. В связи с конвергенцией многих афферентов интернейроны спинного мозга, как и зависимые от них мотонейроны, участвуют не только в замыкании сгибательных и разгибательных рефлексов, но вовлекаются в осуществление двигательных рефлексов ствола и выполнение произвольных движений (рис. 4.21).
4.3.2.5. Ритмические рефлексы
Ритмический чесательный рефлекс вызывается слабым раздражением кожи и включает два компонента: 1) обнаружение раздраженного участка (ползающие насекомые, зуд) и 2) ритмичные чесательные движения, заключающиеся в повторных сгибаниях-разгибаниях конечности, которая занимает при этом определенное положение по отношению к кожной поверхности за счет повышения тонуса приводящих мышц. Комплекс подобных движений направлен на устранение нежелательных ощущений. Чесательный рефлекс сохраняется у собак с изолированным от головного спинным мозгом, что свидетельствует о спинальном механизме этого рефлекса и его относительной независимости от супраспинальных центров.
Правильные чередования активности сгибателей и разгибателей при осуществлении ритмических рефлексов обеспечиваются с помощью интернейронов, которые поочередно тормозят мотонейроны то сгибателей, то разгибателей (рис. 4.22). Чередуемые движения регулирует двигательный центр спинного мозга, составленный из двух «полуцентров»: сгибательного и разгибательного, которые реципрокно тормозят друг друга. Продолжительность торможения в каждом выполняемом цикле зависит от свойств самих тормозных синапсов, в которых происходит динамическое повышение порога возбуждения постсинаптических нейронов.
4.3.2.6. Участие спинного мозга в локомоции
Под локомоцией понимают совокупность координированных движений, с помощью которых человек активно перемещается в пространстве. Его шаг представляет собой цикл движений, в котором участвуют обе конечности, и в каждой из них совершаются ритмические сокращения мышц-сгибателей и разгибателей. Выполняемые каждой ногой попеременно движения представляют собой чередование двух фаз: 1) мах, при котором нога сгибается и отрывается от пола; 2) упор, при котором нога касается пола и разгибается. При этом движения ног синхронизированы так, что одна из них находится в фазе маха, а другая — упора.
 |

 |
|
 |
|
 |
|
 |
Активность мотонейронов, иннервирующих мышцы-антагонисты, проявляется попеременно благодаря включению в рефлекторную дугу тормозных интернейронов, обеспечивающих реципрокное торможение.
В управлении координированной деятельностью мышц при ходьбе и беге участвуют интернейроны и мотонейроны, находящихся в разных сегментах спинного мозга. Согласованное чередование их циклической активности обеспечивается топографически упорядоченными связями между собой, сформировавшимися еще в процессе развития мозга и позволяющими реализовать врожденную программу движений. Спинномозговые центры локомоции контролируются двигательными ядрами ствола, моторной корой и мозжечком, для осуществления контроля используется сенсорная информация (проприоцептивная, вестибулярная, зрительная), переработка которой происходит в головном мозге. Активность спинальных локомоторных центров у человека полностью подчинена супраспинальным отделам и самостоятельно не проявляется.
4.3.2.7. Спинальные вегетативные рефлексы
В зависимости от локализации рецепторов и расположения рабочих органов различают висцеро-висцеральные, висцеродермальные и дермовисцеральные рефлексы. Рефлекторные дуги всех этих рефлексов относятся к поли- синаптическим, рефлекторные реакции происходят при участии гладких мышц внутренних органов и кровеносных сосудов, сердечной мышцы и желез внешней секреции, которые реагируют на раздражение интероре- цепторов или рецепторов кожи.
Висцеро-висцеральные рефлексы вызываются раздражением рецепторов внутренних органов и проявляются изменениями деятельности этих же (например, кардио-кардиальные, кишечно-кишечные, эвакуаторные рефлексы) или других внутренних органов. Висцеродермальные рефлексы возникают при раздражении рецепторов внутренних органов и проявляются изменениями кожной чувствительности, потоотделения и электрического сопротивления кожи. Область рефлекторного ответа определяется сегментарной организацией вегетативной иннервации: афферентные сигналы от рецепторов внутренних органов поступают в определенные сегменты спинного мозга, а кожные изменения обнаруживаются только в тех дерма- | томах, которые иннервируются из этих же сегментов. Изменения кожной > чувствительности или так называемая отраженная боль в ограниченных • участках кожи, получивших название зон Захарьина—Геда, указывают на I поражение определенного органа и имеют диагностическое значение.
Ц Сегментарная организация рефлекторных дуг характерна и для дермо- | висцеральных рефлексов, которые вызываются раздражением рецепторов кожи, после чего происходит рефлекторное изменение деятельности опре- I деленных внутренних органов и возникают реакции сосудов. На сегментарном принципе организации рефлекторных дуг дерматовисцеральных рефлексов основано терапевтическое действие согревания, охлаждения или ■ иного раздражения кожи (точечный массаж, акупунктура, горчичники).
Нисходящий контроль рефлекторных дуг спинальных вегетативных рефлексов осуществляют вегетативные центры ствола мозга и гипоталамус. Окончания серотонинергических, норадренергических и пептидергических нейронов головного мозга проецируются на преганглионарные нейроны и интернейроны, участвующие в замыкании рефлекторных дуг вегетативных ' рефлексов.
4.3.3. Функциональная организация проводящих путей спинного мозга
Проводящие пути спинного мозга образованы топографически упорядоченными пучками длинных аксонов нейронов, входящих в состав задних, передних и боковых канатиков белого вещества спинного мозга. В соответствии с источником и адресатом передаваемой информации они подразделяются на восходящие (чувствительные), нисходящие (двигательные) и собственные, или проприоспинальные, проводящие пути. Собственные проводящие пути образованы аксонами интернейронов и служат для установления межсегментарных связей в пределах самого спинного мозга.
Восходящие пути задних столбов образованы длинными ветвями аксонов чувствительных нейронов спинальных ганглиев, которые следуют к головному мозгу без перекреста и оканчиваются на нейронах ядер задних столбов продолговатого мозга. Они передают сенсорные сигналы от мышечных и сухожильных проприоцепторов и от тактильных рецепторов кожи. Восходящие пути болевой и температурной чувствительности образованы аксонами нейронов задних рогов спинного мозга, которые возбуждаются соответствующими сенсорными нейронами. Аксоны клеток задних рогов проходят через переднюю комиссуру на противоположную сторону и направляются к головному мозгу в составе переднебокового канатика, в котором различают спиноталамический и спиноретикулярный тракты. Спиноталамический тракт служит для проведения сенсорных сигналов от рецепторов в проекционную область коры, а спиноретикулярный тракт проводит нервные импульсы от рецепторов в ретикулярную формацию, которая неспецифически активирует обширные регионы коры. Дорсальный и вентральный спиномозжечковые тракты, расположенные в боковых столбах, передают информацию от механорецепторов кожи, мышц и суставов к мозжечку: это канал обратной связи, необходимой мозжечку для коррекции совершаемых движений.
Нисходящие пути спинного мозга, вестибулоспинальный, руброспиналь- ный и ретикулоспинальный, предназначены для регуляции тонуса скелетных мышц и координации рефлекторной деятельности спинного мозга. Они образованы аксонами нейронов вестибулярного, красного ядер и ретикулярной формации. Берущий свое начало от нейронов покрышки среднего мозга тектоспинальный путь служит для перераспределения тонуса скелетных мышц при совершении ответных реакций на зрительные и слуховые раздражители. Кортикоспинальный путь берет начало от нейронов моторных областей коры, он предназначен для регуляции произвольных движений.
Функции ствола мозга
К стволу мозга относят продолговатый мозг, мост и средний мозг, эти три отдела мозга функционально связаны друг с другом. Ствол содержит ядра II — XII черепных нервов (табл. 4.1), которые обеспечивают афферентную и эфферентную иннервацию тканей головы и внутренних органов, в нем находятся нервные центры двигательных и вегетативных рефлексов. Нервные центры ствола программируют циклическую деятельность (ходьба, жевание, дыхание). Через ствол проходят сенсорные и двигательные пути, обеспечивающие взаимодействие головного и спинного мозга, переключательные ядра ствола являются обязательным звеном для многих из них. Ствол мозга содержит популяции нейронов, которые образуют восходящие и нисходящие проекции, оказывающие модулирующее влияние на осуществление рефлекторной деятельности, поведенческих реакций и психических процессов.
4.4.1. Функциональная организация ствола мозга
Афферентные сигналы поступают к чувствительным ядрам ствола от сенсорных нейронов, тела которых расположены в чувствительных узлах черепно-мозговых нервов. Афферентные волокна образуют синапсы с интернейронами чувствительных ядер, аксоны которых проецируются на двигательные или вегетативные ядра ствола. Эти ядра образованы эфферентными нейронами, их аксоны иннервируют скелетные и гладкие мышцы головы, шеи и внутренних органов, экзокринные железы, которые являются рабочими органами для стволовых рефлексов. В замыкании большинства рефлекторных дуг принимает участие ретикулярная формация, занимающая промежуточное положение между расположенными дорсально чувствительными ядрами и находящимися вентрально двигательными ядрами.
В стволе мозга расположены сегментарные нервные центры собственных рефлексов, которые возникают при активации чувствительных волокон черепно-мозговых нервов и проявляются ответными сокращениями различных групп мышц головы или вегетативными реакциями (например, слюноотделительный, зрачковый рефлексы). Наряду с ними в стволе имеются координационные центры системных рефлексов, возникающих в результате возбуждения чувствительных волокон черепно-мозговых нервов, затем их ядер, от которых оно распространяется по восходящим или нисходящим путям, приводя к цепи моторных и вегетативных рефлексов ствола. Системные рефлексы могут проявляться последовательными сокращениями мышц языка, глотки, гортани, секреторными реакциями, изменениями частоты сокращений сердца, величины артериального давления, частоты дыхания (например, глотание, кашель).
Комплексы ответных рефлекторных реакций на сенсорный стимул, характеризующиеся запрограммированными (автоматическими) действиями, определяются как центральные программы, например программа ходьбы, жевания, дыхания. Под программой понимают заданную генетически последовательность активации нервных центров, которая вызвана пусковым влиянием какого-либо раздражителя. Запущенная центральная программа реализуется автоматически, без дополнительного участия внешних стимулов, однако порядок запрограммированных действий может изменяться в зависимости от характера сенсорной информации (обратная связь).
4.4.1.1. Черепные нервы
В соответствии с основной выполняемой функцией выделяют группу высоко специфических сенсорных черепно-мозговых нервов (И пара — зрительный нерв, VIII пары — слуховой и вестибулярный нервы, I пара — обонятельный нерв). Другую группу представляют смешанные черепно-мозговые нервы, образованные соматическими и висцеральными афферентами и эфферентами: V пара — тройничный нерв, VII пара — лицевой нерв, IX пара — языкоглоточный нерв, X пара — блуждающий нерв. В третью группу включают черепно-мозговые нервы, иннервирующие мышцы головы, в составе которых, помимо аксонов мотонейронов, содержится небольшое количество афферентных волокон от проприоцепторов соответствующих мышц.
4.4.1.2. Функциональная специализация ядер ствола
В ядрах задних столбов продолговатого мозга происходит переключение восходящих сенсорных путей тактильной и проприоцептивной чувствительности. Аксоны получивших афферентную информацию нейронов ядер задних столбов переходят на противоположную сторону, образуя характерные симметричные структуры продолговатого мозга, которые называются лемнисками (от лат. lemnisci — ленты для почетных венков). От этого термина происходит название лемнисковой системы, предназначенной для передачи специфической информации соматосенсорной коре.
Восходящие пути температурной и болевой чувствительности относятся к экстралемнисковой системе, они проходят через ствол без перекрещивания к таламусу, отдавая возбуждающие коллатерали к нейронам ретикулярной формации ствола. Все виды сенсорной информации от специальных органов чувств, за исключением обоняния, проецируются на соответствующие ядра ствола, нейроны которых перерабатывают полученную информацию и передают ее следующему иерархическому уровню {проводниковая функция), представленному ядрами таламуса. Помимо этого афферентные сигналы используются для осуществления собственных или системных рефлексов ствола {рефлекторная функция).
Моторные ядра черепных нервов участвуют в осуществлении рефлекторной деятельности ствола, но в то же время находятся под контролем
Таблица 4.1. Функциональная специализация черепных нервов
| Пара | Название | Афферентный, эфферентный | Специфическая функция |
| 1 | Обонятельный | Афферентный | Обоняние |
| II | Зрительный | Афферентный | Зрение |
| III | Глазодвигательный | Эфферентный Афферентный | Движения глаз, сужение зрачка, изменение формы хрусталика Передача информации от глазодвигательных мышц |
| IV | Блоковый | Эфферентный Афферентный | Движения глаз Передача информации от глазодвигательных мышц |
| V | Тройничный | Эфферентный Афферентный | Жевание Чувствительность лица |
| VI | Отводящий | Эфферентный Афферентный | Движения глаз Передача информации от глазодвигательных мышц |
| VII | Лицевой | Эфферентный Афферентный | Движения мимических мышц лица, выделение слюны и слез Вкусовая чувствительность передних 2/3 языка |
| VIII | Слуховой Вестибулярный | Афферентный Афферентный | Слух Чувство равновесия |
| IX | Языкоглоточный | Афферентный Эфферентный | Вкусовая чувствительность задней 1/3 языка, неба, сведения о газовом составе артериальной крови Выделение слюны, глотание |
| X | Блуждающий | Афферентный Эфферентный | Передача сигналов от внутренних органов Регуляция сокращений гладких мышц, сердца; секреция экзокринных желез |
| XI | Добавочный | Эфферентный Афферентный | Движения головы, шеи, плеч Передача информации от мышц шеи |
| XII | Подъязычный | Эфферентный Афферентный | Движения языка Передача информации от мышц языка |
двигательных областей коры. Берущий начало в этих областях кортикобульбарный тракт регулирует выходную активность двигательных ядер черепно-мозговых нервов и участвует в управлении сокращениями лицевой мускулатуры. Волокна кортикобульбарного тракта оканчиваются также на нейронах координационных центров ствола: красного ядра, вестибулярных ядер, ядер шва, голубого пятна. Эти ядра образуют нисходящие проекции к интернейронам и мотонейронам спинного мозга, посредством которых обеспечивают контроль рефлекторной деятельности спинного мозга и регулируют распределение тонуса скелетных мышц, необходимого для поддержания позы и выполнения движений. Красное и вестибулярное ядра, ядра шва и голубого пятна получают также коллатерали волокон кортикоспинального тракта — прямого пути от двигательной коры к спинному мозгу. Таким образом, моторные ядра ствола участвуют в его собственной рефлекторной деятельности и одновременно являются иерархическим уровнем моторной системы мозга: двигательная кора—ствол—спинной мозг.
При осуществлении двигательной функции ЦНС ядра ствола взаимодействуют с мозжечком\ нейроны таких ядер активируются сенсорными системами (вестибулярная, проприоцептивная, зрительная), возбуждающимися в процессе движения, а затем передают информацию мозжечку, который, в свою очередь, регулирует выходную активность координационных центров ствола, в первую очередь, вестибулярных ядер. У мозжечка нет прямых связей с корой, и все его афферентные и эфферентные связи с ней осуществляются посредством нескольких ядер ствола (ретикулярные ядра продолговатого мозга, ядра нижней оливы и моста).
В регуляции жизненно важных вегетативных функций (кровообращение, дыхание, пищеварение) участвуют относительно небольшие, сложно организованные группы нейронов ретикулярной формации, координирующие активность симпатического и парасимпатического отделов вегетативной нервной системы. Большинство нейронов ретикулярной формации являются полимодальными\ они получают разные виды сенсорной информации, а также проекции нейронов гипоталамуса и лимбической системы мозга. Гипоталамус координирует деятельность вегетативных центров ствола, а лимбическая система изменяет их активность при возникновении эмоций.
Компактно сгруппированные в нескольких ядрах ствола моноаминерги- ческие нейроны (ядра шва, голубого пятна, среднего мозга) образуют дивергентные восходящие и нисходящие проекции, способные повлиять на процесс передачи нервных импульсов в различных регионах мозга (таламус, лимбические структуры, полосатое тело, кора). Нейромодулирующее действие моноаминергических нейронов используется при регуляции циклических процессов (чередование сна и бодрствования, распределение внимания), возникновении мотиваций и эмоций.
4.4.2. Рефлекторная функция ствола мозга
Рефлексы ствола участвуют в осуществлении функций пищеварения (выделение слюны, жевание, глотание), защиты от повреждающих воздействий (рвота, чиханье, кашель, слезоотделение, мигание) и приспособлении к меняющимся условиям среды (ориентировочные, сторожевые рефлексы).
Слюноотделительный рефлекс возникает при раздражении вкусовых рецепторов и механорецепторов полости рта, афферентная импульсация проводится по чувствительным волокнам лицевого и языкоглоточного, а также тройничного и блуждающего нервов. Слюноотделительный центр продолговатого мозга представляет функциональное объединение нейронов, расположенных в ретикулярной формации, которые активируются при раздражении вкусовых и обонятельных рецепторов. Это координационный центр, который связан не с одним, а с несколькими вегетативными ядрами. В частности, рефлекторное выделение слюны происходит в результате одновременной активации парасимпатических волокон лицевого и языкоглоточного нервов, а также симпатических нейронов из II—IV грудных сегментов спинного мозга. Стимуляция слюноотделения одним лишь запахом, видом пищи или воспоминаниями о ней указывает на возможность активации слюноотделительного центра нисходящими корковыми проекциями.
Рецептивные поля глотательного рефлекса находятся в корне языка, небе, гортани и надгортаннике, где расположены механорецепторы тройничного и языкоглоточного нервов, а также верхнегортанной ветви блуждающего нерва. Они раздражаются пищевым комком, проглатываемой жидкостью или слюной, после чего афферентная импульсация поступает к ядрам тройничного и солитарного трактов и достигает глотательного центра ретикулярной формации, находящегося между ядром лицевого нерва и нижней оливой. Эфферентная импульсация, обеспечивающая координированные сокращения мышц рта, глотки и пищевода, поступает к ним по волокнам подъязычного и лицевого нервов, а также при посредстве мотонейронов первых трех шейных сегментов спинного мозга. Глотательный центр функционально связан с дыхательным центром, эта связь обеспечивает задержку дыхания при глотании и перекрытие надгортанником входа в дыхательные пути во избежание поступления в них пищи.
Жевательные рефлексы (открывания и закрывания рта) осуществляются в соответствии с центральной программой жевания. Она запускается стимуляцией механорецепторов при изменениях давления на твердое небо, зубы, десны. Афферентная информация поступает в продолговатый мозг по чувствительным волокнам тройничного нерва, двигательное ядро которого осуществляет эфферентную иннервацию жевательных мышц. Представление о жевании как о рефлексе описывает этот процесс в виде чередования двух реципрокных фаз: открывания и закрывания рта. В современном представлении центр жевания функционирует как центральный генератор ритмических мышечных сокращений, который зависит от сенсорных сигналов (поступление твердой пищи в ротовую полость) и от влияния центральных структур (жевание можно начать и прекратить произвольно). Нисходящий контроль центра жевания осуществляют двигательные ядра ствола, моторные области коры, мозжечок и базальные ганглии.
Рвотный рефлекс является результатом раздражения механо- и хеморецепторов, расположенных в корне языка, глотке, слизистой оболочке желудка, кишечника, брюшины, а также стимуляцией обонятельных, вкусовых рецепторов (при резко неприятном запахе или вкусе пищи) и вестибулярных рецепторов внутреннего уха (вследствие укачивания). Афферентные импульсы поступают в ретикулярную формацию продолговатого мозга по волокнам блуждающего, языкоглоточного и других чувствительных нервов. Рвотный центр осуществляет сложную координацию соматических и висцеральных двигательных реакций, приводящих к извержению содержимого желудка, а иногда и кишечника, через рот. При расслаблении дна желудка и сфинктера между желудком и пищеводом происходит непроизвольное сокращение диафрагмы и брюшной стенки одновременно с сокращением привратника, что приводит к выталкиванию содержимого желудка в пищевод. Повышение внутриплеврального давления способствует продвижению содержимого желудка из пищевода в ротовую полость. Рефлекторный подъем мягкого неба предупреждает попадание рвотных масс в носовую часть глотки, а рефлекторное закрытие голосовой щели и прекращение дыхания предотвращают аспирацию содержимого желудка в легкие. Эфферентные пути рвотного рефлекса представлены волокнами блуждающего и языкоглоточного нервов, а также мотонейронами, иннервирующими мышцы брюшной стенки и диафрагму. Рвотный центр находится под контролем кортикальных центров мозга.
Чихательный рефлекс обеспечивает удаление инородных веществ, попавших в полость носа, и возникает при механическом раздражении чувствительных окончаний тройничного нерва. Сначала происходит глубокий вдох, а затем форсированный выдох при открытой голосовой щели. Поднимающийся к небу язык закрывает путь в ротовую полость, и поток выходящего воздуха идет через нос, освобождая его полость от инородных частиц. Чихательный рефлекс представляет собой сложный моторный акт с жестко запрограммированной последовательностью действий, которой управляет центральный генератор, связанный с дыхательным центром. Возбуждение чувствительных волокон передается нейронам дыхательного центра и эфферентным волокнам языкоглоточного, блуждающего, подъязычного нервов; в акте чиханья участвуют и мотонейроны дыхательных мышц. Запущенный механизм чихательного рефлекса всегда приводит к его завершению вне зависимости от поступления сенсорных сигналов.
Кашлевой рефлекс возникает при раздражении находящихся в носоглотке, гортани и трахее чувствительных окончаний тройничного, языкоглоточного, верхнего гортанного и блуждающего нервов. Механическими раздражителями этих чувствительных окончаний являются вдыхаемые с воздухом твердые частицы или продукты воспалительных процессов, химическими раздражителями служат вдыхаемые газы. Вследствие раздражения чувствительных окончаний происходит глубокий вдох, закрывается голосовая щель, расслабляется диафрагма, а скелетные дыхательные мышцы сокращаются, что в итоге приводит к повышению внутригрудного давления, которому противодействует закрытая голосовая щель. При открытии голосовой щели возникает быстрый выдох, способствующий удалению инородных тел или воспалительной слизи. При кашлевом рефлексе происходит согласование активности дыхательного центра с нейронами, управляющими деятельностью мышц гортани и скелетных мышц. Эфферентное звено рефлекса включает возвратный гортанный нерв, регулирующий закрытие голосовой щели, и спинномозговые нервы, контролирующие сокращения грудных и брюшных мышц.
Слезный рефлекс обусловлен подсыханием роговицы или попаданием на нее инородных тел, рефлекторный механизм защищает роговицу и конъюнктиву от высыхания и от раздражения инородными телами, смываемыми слезной жидкостью. Афферентная импульсация от роговицы осуществляется по волокнам слезной ветви тройничного нерва, в переключении сигналов, следующих к слезным железам, участвуют чувствительные ядра тройничного тракта и ретикулярная формация. Эфферентное звено представляют парасимпатические нейроны, стимулирующие выделение слезной жидкости. Преганглионарные парасимпатические нейроны расположены в области моста, они могут возбуждаться и вызывать повышенное выделение слезной жидкости при чиханье, кашле, рвоте, жевании, действии яркого света, а также при переживании эмоций. Сопутствующее указанным реакциям слезотечение объясняется стимуляцией парасимпатических нейронов из чувствительного ядра тройничного нерва, гипоталамуса и лимбической системы. Активация симпатического отдела вегетативной нервной системы приводит к уменьшению количества выделяемой слезной жидкости.
При раздражении роговицы помимо слезного возникает мигательный рефлекс, связанный с раздражением чувствительных окончаний тройничного нерва. Рефлекторное опускание века происходит вследствие сокращения орбикулярной мышцы, получающей иннервацию от лицевого нерва. При движении века инородное тело удаляется с роговицы и уносится слезной жидкостью, а сама слезная жидкость равномерно распределяется по роговице и конъюнктиве.
В верхнем отделе ствола, представленном структурами среднего мозга, расположен центр зрачкового рефлекса, который служит для регуляции потока света, поступающего в глаз: на ярком свету зрачок суживается, а в темноте расширяется. Афферентные импульсы от сетчатки глаза поступают к верхним буграм четверохолмия, где находятся нейроны, осуществляющие замыкание рефлекторной дуги в зависимости от характера афферентной импульсации. При ярком освещении центр зрачкового рефлекса активирует вегетативные ядра глазодвигательных нервов, которые иннервируют мышцы, суживающие зрачок. В темноте характер афферентной импульсации изменяется, в связи с этим рефлекторная дуга замыкается на симпатические нейроны, находящиеся в нижнем шейном и верхнем грудном сегментах спинного мозга. Происходит активация этих нейронов, что ведет к сокращению расширяющих зрачок радиальных мышц.
Если верхние (передние) бугры четверохолмия получают афферентные сигналы от сетчатки, то нижние (задние) — от слуховых рецепторов. При действии новых зрительных или слуховых раздражителей здесь замыкаются рефлекторные дуги ориентировочных рефлексов, проявляющихся поворотом головы и глаз в сторону раздражителя и одновременным перераспределением тонуса скелетных мышц так, чтобы получилась поза, из которой легче начать движение. Указанные реакции получили название сторожевых или стартовых рефлексов. Физиологическая основа их возникновения описана в следующих разделах этой главы.
4.4.2.1. Статические и статокинетические рефлексы
Статические и статокинетические рефлексы обеспечивают степень тонического напряжения мышц, фиксирующих неподвижное положение суставов, которое необходимо для поддержания позы и сохранения равновесия и ориентации конечностей во время движения. Статические рефлексы подразделяются на позные, или рефлексы положения, благодаря которым сохраняется вертикальная поза, и установочные (выпрямления) рефлексы, проявляющиеся при смене одной позы на другую, например при вставании из положения сидя или лежа. Статокинетические рефлексы вызываются действием на организм прямолинейного или углового ускорения.
Одним из источников афферентной импульсации, требующейся для возникновения обеих разновидностей рефлексов, служат рецепторы вестибулярного аппарата, реагирующие на изменения положения тела, наклоны и повороты головы. Еще одну рефлексогенную зону образуют проприоцепторы шейных мышц, возбуждающиеся в связи с наклонами головы. В соответствии с источником афферентной импульсации различают вестибулярные (или лабиринтные) и шейные тонические рефлексы. Определение рефлексов как тонических указывает на перераспределение тонуса мышц, требуемого для сохранения равновесия и необходимой позы при смещении
| Шея | Лабиринт | ||
| голова вверх | нормальное положение головы | голова вниз | |
| Отклонение дорсально | А | Б | в |
| Нормальное отклонение |
| д | Е |
| Отклонение вентрально | У | 3 | И |
Рис. 4.23. Схема лабиринтных и шейных рефлексов: распределение тонуса мышц конечностей.
Тонус мышц, сгибающих или разгибающих конечности, изменяется рефлекторно в соответствии с положением головы относительно горизонта (афферентные сигналы поступают от вес- тибулорецепторов) и наклонами головы (афферентные сигналы поступают от проприоцепторов мышц шеи). Окончательный вариант распределения тонуса мышц на конечностях зависит от характера афферентной информации, поступившей из обоих источников.
центра тяжести тела. Для удержания равновесия необходимо повысить тонус мышц, противодействующих силе гравитации. К таким мышцам относятся разгибатели туловища и проксимальных отделов конечностей (рис. 4.23).
Рефлекторные дуги вестибулярных и шейных рефлексов замыкаются в соответствующих сенсорных ядрах продолговатого мозга, нейроны которых образуют проекции на стволовые центры нисходящих двигательных путей, оканчивающихся в сером веществе спинного мозга. Двигательные центры ствола представлены крупными нейронами красного ядра (его маг- ноцеллюлярная часть), вестибулярными ядрами, медиальной частью ретикулярной формации и покрышкой среднего мозга. Нейроны этих ядер образуют нисходящие проекции на интернейроны спинного мозга и гамма-мотонейроны, что позволяет им координировать активность мотонейронов, непосредственно управляющих мышечными сокращениями.
Позные рефлексы проявляются при подготовке к любому движению, поскольку для его выполнения требуется определенная исходная поза: например, для того, чтобы встать из положения сидя, вначале необходимо чуть наклонить вперед корпус и голову. Подъем с постели тоже начинается с опережающего изменения положения головы, при котором возбуждаются вестибулярные рецепторы, проприоцепторы шеи и происходит рефлекторное перераспределение тонуса мышц туловища и конечностей, необходимое для того, чтобы подняться.
Примером статокинетического рефлекса может служить сохранение равновесия у стоящего в транспорте пассажира, когда это равновесие нарушается при резком начале движения или внезапной остановке. При смещении центра тяжести рефлекторно повышается тонус разгибателей на той стороне, в которую отклоняется тело, а выставленная рефлекторно в эту сторону прямая нога помогает удержать равновесие. При вертикальных ускорениях имеют место лифтные рефлексы', в момент начала движения лифта вверх у находящегося на лифтной площадке человека снижается тонус разгибателей, и поэтому его ноги подгибаются, а при опускании площадки тонус разгибателей увеличивается и ноги фиксируются в положении максимального разгибания.
Во время ходьбы и бега возникает смещение центра тяжести тела вперед. Если его уже нельзя вернуть в исходное положение, не оторвав конечность от опоры, то для сохранения равновесия приходится сделать шаг вперед. В том случае, если поскользнувшийся человек начинает падать, на стороне падения рефлекторно повышается тонус разгибателей. Этот эволюционно древний рефлекторный механизм приводит к повышению тонуса разгибателей не только ноги, выставленной в сторону падения, но и руки, что нередко приводит к типичному перелому лучевой кости, когда при падении на нее приходится вся сила удара. Спортсмены, чья деятельность предполагает частые падения, учатся это делать безопасно, а овладение новой техникой указывает на возможность перепрограммирования статокинетического рефлекса, что связано с участием двигательных центров мозжечка и моторных областей коры.
В клинической практике статические и статокинетические рефлексы называют постуральными и исследуют их электромиографически.
4.4.2.2. Нисходящие двигательные пути ствола мозга
Связь двигательных центров ствола с интернейронами и мотонейронами спинного мозга обеспечивается двумя различающимися в функциональном отношении нисходящими путями: медиальным и латеральным (рис. 4.24). Медиальный путь образуют волокна вестибулоспинального, ретикулоспинального и тектоспинального трактов, которые оканчиваются билатерально на медиальных колоннах вентральных рогов спинного мозга, где находятся интернейроны и мотонейроны, управляющие мышцами туловища и проксимальных отделов конечностей. Основная функциональная задача этого пути состоит в управлении деятельностью мышц-разгибателей, обеспечивающих сохранение равновесия и вертикального положения тела. Еще одна его функция заключается в модулировании активности возбуждающих и тормозных интернейронов спинного мозга, что проявляется торможением или облегчением спинальных рефлексов. Специфической задачей текто-
 |
|
 |

А. Латеральный путь начинается от крупноклеточных нейронов красного ядра и оканчивается на противоположной стороне спинного мозга.
Б. Медиальный путь образован аксонами нейронов вестибулярного ядра, ретикулярной формации и покрышки среднего мозга; в спинном мозге представлен передним канатиком, оканчивается билатерально.
спинального тракта является координация поворотов головы и следящих движений глаз с изменениями позы.
Латеральный путь от ствола к спинному мозгу представлен руброспи- налъным трактом, волокна которого переходят на противоположную сторону и после прохождения ствола спускаются в спинной мозг, где образуют синапсы с интернейронами и мотонейронами, расположенными лате- рально и контролирующими деятельность дистальных мышц конечностей. Эти мышцы предназначены для выполнения сложных движений, связанных, например, с захватом предметов рукой и пальцами.
После перерезки руброспинального пути у экспериментальных животных возникает стойкое повышение тонуса разгибателей туловища и конечностей, что получило название децеребрационной ригидности. Она исчезает после дополнительной перерезки вестибулоспинального и ретикулоспинального путей. Сопоставление этих фактов дало повод к выводу о необходимости красных ядер для противодействия стволовым антигравитационным центрам, т. е. вестибулярным ядрам и ретикулярной формации. Однако у человека после изолированного поражения красного ядра повышение тонуса разгибателей выражено Незначительно, а иногда повышения их тонуса и вовсе не наблюдается. Только при массивных очагах повреждения в области верхнего двухолмия, когда страдает и кортикоспинальный путь, обнаруживается повышенный тонус разгибателей в нижних конечностях, но одновременно с этим в верхних конечностях преобладает тонус сгибателей. Возможно, это обстоятельство обусловлено изменением функционального назначения верхних конечностей у человека по сравнению с четвероногими.
4.4.2.3. Глазодвигательные центры ствола
Движения глазного яблока осуществляют четыре прямые мышцы (наружная, внутренняя, верхняя, нижняя) и две косые мышцы (верхняя и нижняя). Они поворачивают глаз внутрь или наружу, вверх или вниз, чтобы сфокусировать рассматриваемые объекты в центральной ямке сетчатки, обеспечивающей максимальную остроту зрения; при этом движения глазных яблок происходят содружественно. Сокращения мышц определяются влиянием глазодвигательных и блоковых нервов, ядра которых расположены в ростральной части среднего мозга, а также ядер отводящих нервов, содержащихся в каудальной части моста. Различают очень быстрые движения глаз {саккады) и плавные следящие движения. Саккады, происходящие около трех раз в секунду, обеспечивают перемещение изображения по рецепторам сетчатки, которые не реагируют на стабильное изображение, а отвечают на его движение. Плавные следящие движения глаз возникают в связи с перемещением наблюдаемых объектов и при движении головы и тела наблюдателя. Следящие движения позволяют удерживать изображение в пределах центральной ямки сетчатки.
Афферентная информация к мотонейронам глазодвигательных мышц поступает от зрительных, вестибулярных, слуховых рецепторов, а также от проприоцепторов мышц глаза. В зависимости от характера афферентной информации дуги глазодвигательных рефлексов замыкаются на разных уровнях ствола. Стволовые центры, осуществляющие замыкание рефлекторных дуг и обеспечивающие определенное положение взора, расположены в парамедианной ретикулярной формации и ретикулярной формации среднего мозга. Уровень возбуждения глазодвигательных центров регулируют нейроны верхних бугров четверохолмия, вестибулярные ядра и мозжечок, кора больших полушарий.








