2020-05-25
2020-05-25 163
163
Сенсорные системы осуществляют непрерывное слежение за состоянием внешней и внутренней среды и немедленно оповещают об изменениях параметров среды. Моторные системы обеспечивают действия, направленные на уравновешивание с меняющимся состоянием среды. Мотивационные системы получают сенсорную информацию, регистрируют прежний опыт приспособления к условиям среды и побуждают к действиям, направленным на удовлетворение возникающих потребностей.
ластям коры, создающим замысел последующих действий. В соответствии с этим замыслом моторные системы формируют программы движений. Само решение о начале, как и об окончании, действий принимает мотивационная система, влияющая на выходную моторную активность, а через гипоталамус - и на состояние вегетативных функций.
Таким образом, сенсорные системы предоставляют информацию об изменениях внешней или внутренней среды, моторные системы обеспечивают ответные действия, направленные на приспособление к изменившимся условиям. Мотивационные системы активируются при возникновении каких-либо потребностей и побуждают к совершению действий, направленных на удовлетворение потребности. Функциональные системы постоянно взаимодействуют друг с другом: сенсорные системы активируют мотивационные, а они побуждают моторные к созданию оптимальных для сложившейся ситуации двигательных программ (рис. 4.9).
4.2.1. Общие принципы организации функциональных систем мозга
В любой из функциональных систем соблюдаются единые принципы ее организации.
4.2.1.1. Существование нескольких уровней переработки информации
Передача сигналов от одних нейронов к другим происходит в переключательных ядрах, находящихся в спинном и головном мозге. Каждое переключательное ядро представляет собой, во-первых, распределительную систему, образующую проекции на другие структуры мозга для передачи им информации. Во-вторых, в любом переключательном ядре происходит переработка информации, заключающаяся в изменениях параметров электрических сигналов. Переключательные ядра образованы разными типами нервных клеток, которые, в зависимости от выполняемой функции, подразделяются на два класса: проекционные и локальные интернейроны.
Проекционные нейроны имеют достаточно длинные аксоны, с помощью которых переносят электрические сигналы от одного переключательного ядра к другому, передавая информацию на следующий уровень ее переработки. Короткие аксоны возбуждающих и тормозных локальных интернейронов не выходят за пределы переключательного ядра. Функция локальных интернейронов состоит в облегчении передачи одних сигналов и подавлении других, что способствует передаче на следующий уровень наиболее значимых сигналов и подавлению «информационного шума».
4.2.1.2. Топографическая упорядоченность проводящих путей
В переключательных ядрах и в проводящих путях сенсорных систем электрические сигналы распространяются упорядоченно от каждого рецептивного поля, например в сетчатке глаза или коже, к соответствующей именно ему проекционной области коры. Благодаря этому проекционная кора представляет собой своего рода нейронную карту, отражающую пространственные характеристики действующих стимулов. Нейроны моторной коры тоже расположены топографически упорядоченно и могут влиять только на иерархически подчиненные им нейроны головного и спинного мозга для передачи команд строго определенным мышцам.
4.2.1.3. Наличие параллельных проводящих путей
В каждой функциональной системе мозга существуют независимые параллельные пути переработки специализированной информации. Например, в зрительной сенсорной системе независимо и параллельно перерабатывается информация о форме, цвете и движении каждого объекта, оказавшегося в зрительном поле. В двигательных системах существуют раздельные специализированные пути, по которым информация поступает от высших двигательных центров к мотонейронам спинного мозга: одни пути служат для контроля положения тела, рефлексов спинного мозга, другие — для управления точными движениями кистей рук и пальцев.
4.2.2. Типы нейронных сетей
Специфические функциональные соединения нервных клеток друг с другом образуют нейронные сети, в зависимости от характера межнейронных соединений различают три типа сетей: иерархические, локальные и дивергентные.
Иерархический тип нейронной сети характерен для сенсорных и моторных систем, он включает несколько уровней переработки информации, представленных переключательными ядрами. В сенсорных системах информация от первичных сенсорных нейронов передается вторичным сенсорным нейронам, от них — нейронам третьего порядка и далее до проекционной зоны коры. В моторных системах существует нисходящая иерархия: созданные в двигательных областях коры команды поступают к переключательным ядрам ствола, оттуда — в спинной мозг, а из него — к мышцам. Передача сигналов в синапсах иерархических сетей происходит с помощью возбуждающих нейромедиаторов. На каждом уровне иерархической организации имеются локальные сети, образованные возбуждающими и тормозными интернейронами. Функция нейронов локальной сети состоит в сужении или расширении передаваемого потока информации и ее контрастировании путем усиления информационно значимых сигналов и подавления шума.
Дивергентные сети образованы небольшими компактными группами нейронов, тела которых находятся в нескольких ядрах ствола, а многочисленные ветви их длинных аксонов распространяются диффузно ко многим структурам мозга, включая кору. Области проекции таких нейронов предназначены для выполнения разных функций, поэтому дивергентные сети принято связывать с неспецифической деятельностью мозга. Наиболее изученные дивергентные сети в головном мозге образованы норадренергическими, серотонинергическими, дофаминергическими и холинергическими нейронами моста, продолговатого и среднего мозга. Действие медиаторов дивергентных систем модулирует передачу сигналов, т. е. облегчает или затрудняет ее, что изменяет характер интегративной деятельности мозга (чередование сна и бодрствования, формирование внимания, возникновение мотиваций и эмоций).
4.2.3. Нейрохимические классы нейронов
Разные популяции нейронов синтезируют различные нейромедиаторы: синтез конкретного медиатора обусловлен имеющимся в клетке набором ферментов, который, в свою очередь, определяется экспрессией тех или иных генов. Многие нейроны используют комбинацию из одного низкомолекулярного и одного или нескольких пептидных нейромедиаторов, способных усилить или ослабить действие низкомолекулярного медиатора. В первом случае они действуют на общую постсинаптическую мишень как синергисты (например, ацетилхолин и вазоинтестинальный пептид), во втором как — антагонисты (глутамат и дайнорфин в гиппокампе, где глутамат возбуждает, а опиоидный пептид дайнорфин уменьшает активность постсинаптической клетки).
Нейроны, синтезирующие определенный медиатор, образуют популяции, распределение которых в разных регионах мозга строго упорядочено. Эта упорядоченность позволяет выделить глутаматергическую, холинергическую. дофаминергическую. ГАМКергическую и т. п. системы нейронов.
4.2.3.1. Глутаматергическая система
Глутамат является медиатором большинства сенсорных нейронов, пирамидных клеток коры, многих нейронов гиппокампа, мозжечка, ствола и спинного мозга. Это самый распространенный в головном мозге возбуждающий медиатор, для которого существуют ионотропные и метаботропные рецепторы. Ионотропные рецепторы подразделяют в зависимости от чувствительности к NMDA (К-метил-1)-аспартату), АМРА (аминометилпропио- нату) и каиновой кислоте (рис. 4.10). Метаботропные глутаматные рецепторы после присоединения к ним медиатора изменяют чувствительность к глутамату у ионотропных рецепторов, расположенных на общей постсинаптической мембране.
Самые быстрые возбуждающие процессы в глутаматергических синапсах происходят в результате активации АМРА-рецепторов. Они содержат
 |

 | | ||||
 | |||||
|
A. Ионотропные АМРА-рецепторы: присоединение глутамата (Глу) приводит к открытию канала для ионов натрия; рецепторы чувствительны к бензодиазепинам (2,3-БДЗ).
Б. Ионотропные NMDA-рецепторы представляют собой белковую молекулу, содержащую канал для ионов кальция, который при значении мембранного потенциала покоя блокирован ионами магния. Для открывания канала с последующим током через него ионов кальция необходимо присоединение глутамата к рецептору и деполяризация мембраны, после которой ионы магния освобождают канал. Рецепторы чувствительны к концентрации глицина (Гли) в окружающей среде.
B. Ионотропные рецепторы для глутамата, одновременно чувствительные к каиновой кислоте (КА); содержат канал для ионов натрия и калия.
Г. Метаботропные рецепторы для глутамата: после присоединения нейромедиатора происходит активация вторичных посредников. Рецепторы чувствительны к квискулату (КВ И С).
катионные каналы для ионов калия и натрия и используются для получения сигналов от афферентных нейронов. Рецепторы NMDA-типа имеют катионные каналы, пригодные для тока не только ионов калия и натрия, но и кальция, однако ток ионов возможен только при условии предварительной деполяризации мембраны, поскольку при значении мембранного потенциала покоя каналы блокированы ионами магния. При достаточно длительной деполяризации постсинаптической мембраны, обусловленной активацией нечувствительных к NMDA рецепторов, ионы магния уступают вход в канал ионам кальция, которые поступают в постсинаптический нейрон.
При низкой активности пресинаптического нейрона и соответственно малом выделении глутамата постсинаптический потенциал возникает преимущественно за счет активации нечувствительных к NMDA ионотропных рецепторов, например АМРА. Если же ритмическая активность пресинаптического нейрона будет продолжаться, то произойдет ток ионов кальция через каналы NMDA рецепторов. Повышение концентрации ионов кальция в постсинаптическом нейроне вызовет активацию нескольких систем вторичных посредников: протеинкиназы С, кальций-кальмодулин-зависи- мой протеинкиназы и тирозинкиназы. Активация вторичных посредников приведет к длительному возбуждению постсинаптического нейрона, сохраняющемуся часами или сутками (долговременная потенциация).
Вторичные посредники обеспечивают открытое состояние каналов NMDA рецепторов, и, кроме того, в ходе происходящих в постсинаптическом нейроне процессов в нем образуются и выделяются оксиды азота и углерода, Эти низкомолекулярные вещества диффундируют к пресинапти- ческому нейрону и вызывают в нем активацию посредников, способствующих длительному выделению нейромедиатора. Длительная, а тем более повторяющаяся долговременная потенциация в одних и тех же синапсах приводит к экспрессии генов и синтезу белков, необходимых для структурной перестройки синапсов. Перестройка заключается в увеличении количества дендритных шипиков постсинаптического нейрона и вступающих с ними в контакт терминальных окончаний пресинаптического нейрона. Эти изменения происходят в синапсах, которые активируются при часто повторяющемся действии одних и тех же внешних раздражителей, а также при совпадении действующих стимулов друг с другом во времени. Возникшие в синапсах преобразования являются материальной основой энграмм (следов) долговременной памяти.
4.2.3.2. Холинергическая система
Ацетилхолин в качестве нейромедиатора используется в ядрах черепно-мозговых нервов, мотонейронах спинного мозга и нейронах вегетативной нервной системы. В головном мозге холинергические нейроны представлены компактными популяциями в базальных ядрах, перегородке и ядрах диагонального пучка Брока. Их аксоны образуют проекции к структурам переднего мозга, гиппокампу и коре, на которые ацетилхолин действует через М~холинорецепторы. Ацетилхолин используется в качестве нейромодулятора, повышающего эффективность переработки сенсорной информации и ассоциативных процессов в коре. Большое число холинергических нейронов имеется в полосатом теле, где они выполняют функцию локальных интернейронов.
Помимо переднего мозга холинергические нейроны имеются в ростральном отделе ствола: они связывают средний мозг с таламусом и корой больших полушарий. Активация этих нейронов у спящих ведет к пробуждению и одновременно обусловливает изменения ритма электроэнцефалограммы, который становится характерным для состояния бодрствования. В коре преобладают холинорецепторы мускаринового типа, а в некоторых подкорковых структурах (голубое пятно, супраоптическое ядро гипоталамуса, отдельные ядра таламуса) существуют Н-холинорецепторы, с участием которых осуществляются быстрые синаптические процессы. Таким образом, ацетилхолин в головном мозге может действовать через мускаринчувствительные рецепторы в качестве нейромодулятора и выполняет функцию классического нейромедиатора при действии на никотинчувствительные рецепторы.
4.2.3.3. Системы нейронов, использующих биогенные амины
К биогенным аминам относят катехоламины {дофамин, норадреналин, адреналин) и серотонин.
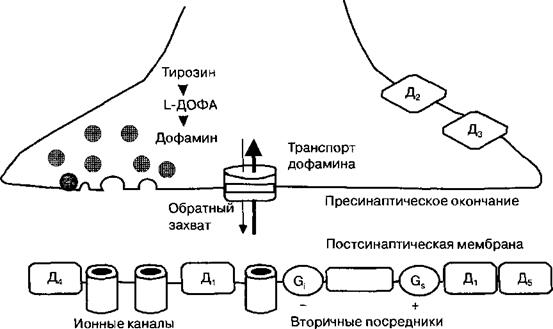
Синтезирующие дофамин нейроны находятся, в основном, в среднем и промежуточном мозге, образуя нигростриатную, мезолимбическую (мезокор- тикальную) и гипоталамо-гипофизарную дофаминэргические системы. Ниг- ростриатная система представлена дофаминергическими нейронами компактной части черной субстанции среднего мозга, их аксоны достигают полосатого тела; они необходимы для осуществления произвольных движений человека. Мезолимбическую (мезокортикальную) систему представляют локализованные вне черной субстанции нейроны среднего мозга, аксоны которых оканчиваются в лимбических структурах мозга (миндалины, перего-