2020-05-25
2020-05-25 267
267В желудке пища, смешанная со слюной и слизью, задерживается от 3 до 10 ч для ее механической и химической обработки. Желудок осуществляет следующие функции: 1) депонирование пищи\ 2) секрецию желудочного сока, обеспечивающего химическую обработку пищи; 3) перемешивание пищи с пищеварительными соками; 4) ее эвакуацию — передвижение порциями в двенадцатиперстную кишку; 5) всасывание в кровь небольшого количества веществ, поступивших с пищей; 6) выделение (экскрецию) вместе с желудочным соком в полость желудка метаболитов (мочевины, мочевой кислоты, креатина, креатинина), веществ, поступивших в организм извне (солей тяжелых металлов, йода, фармакологических препаратов); 7) образование активных веществ (инкрецию), принимающих участие в регуляции деятельности желудочных и других пищеварительных желез (гастрина, гистамина, соматостатина, мотилина и др.); 8) бактерицидное и бактериостатическое действие желудочного сока; 9) удаление недоброкачественной пищи, предупреждающее ее попадание в кишечник.
11.5.1. Секреторная функция желудка
Секреторная функция желудка осуществляется желудочными железами, продуцирующими желудочный сок. Они состоят из трех видов клеток: главных, принимающих участие в выработке ферментов; обкладочных (париетальных), участвующих в выработке хлористоводородной (соляной) кислоты, и добавочных, выделяющих мукоидный секрет (слизь). В его состав входит также внутренний фактор кастла (гастромукопротен), участвующий в регуляции кровотворения. Натощак слизь выделяется также цилиндрическим эпителием, которым покрыта слизистая оболочка желудка. Железы кардиального отдела желудка секретируют в основном слизь. В железах пилорического отдела отсутствуют обкладочные клетки. Поэтому в секрете желез этого отдела отсутствует соляная кислота и его pH равен 7,8—8,4. Основную роль в желудочном пищеварении играют железы фундального отдела, включающего три секректорные зоны: дна, малой кривизны и тела желудка (рис. 11.11). Эти железы имеют все три типа клеток и выделяют основное количество желудочного сока.
Состав желудочного сока. В состоянии покоя (натощак) из желудка человека можно извлечь около 50 мл желудочного содержимого нейтральной или слабокислой реакции (pH 6,0). Это смесь слюны и желудочного сока.
Общее количество желудочного сока, отделяющегося у человека при обычном пищевом режиме, составляет 2,0—2,5 л в сутки. Это бесцветная,
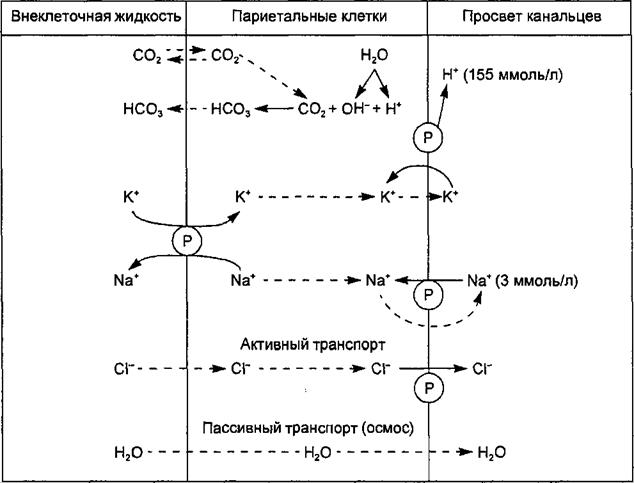
Рис. 11.11. Образование соляной кислоты желудочного сока. Пояснения в тексте. Символ ® означает активность ферментных транспортных систем мембраны кислотопродуцирующих клеток. Стрелками показано направление движения ионов и воды.
прозрачная, слегка опалесцирующая жидкость с удельным весом 1,002— 1,007. В соке могут быть хлопья слизи. Желудочный сок имеет кислую реакцию (pH 0,8—1,5) вследствие высокого содержания в нем хлористоводородной (соляной) кислоты (0,3—0,5 %). Содержание воды в соке — 99,0— 99,5 %, а плотных веществ — 1,0—0,5 %. Плотный остаток представлен органическими и неорганическими веществами: хлоридами (5—6 г/л), сульфатами (10 мг/л), фосфатами (10—60 мг/л), гидрокарбонатами (0—1,2 г/л) натрия, калия, кальция и магния, аммиалом (20—80 мг/л). Значительная часть минеральных веществ всасывается в желудке и кишечнике в кровь и участвует в поддержании постоянства внутренней среды. Основной неорганический компонент желудочного сока — соляная кислота (см. ниже). Органическая часть плотного остатка состоит из ферментов и мукоидов (см. ниже). В небольшом количестве находятся в остатке азотсодержащие вещества небелковой природы (мочевина, мочевая кислота, молочная кислота и др.), подлежащие удалению из организма.
Механизм секреции соляной кислоты. Хлористоводородная кислота вырабатывается париетальными (обкладочными) клетками желез желудка. Эти клетки характеризуются богатством митохондрий, расположенных вдоль внутриклеточных канальцев. Площадь мембраны канальцев и апикальной поверхности клеток во время стимуляции на высоте секреции резко возрастает за счет встроенных в мембрану тубовезикул (трубочек-пузырьков), что сопровождается значительным увеличением клеточных канальцев, проникающих вплоть до базальной мембраны. Это значительно увеличивает возможности синтеза гландулоцитом соляной кислоты. Вдоль канальцев располагается множество митохондрий, площадь внутренней мембраны которых возрастает в процессе биосинтеза НС1. Соответственно увеличивается площадь контакта канальцев и апикальной мембраны клетки. Таким образом, увеличение секреторной активности париетальных клеток обусловлено увеличением площади секреторной мембраны.
Секреция НО является ярко выраженным цАМФ-зависимым процессом, активация которого протекает на фоне усиления гликогенолитической и гликолитической активности, что сопровождается продукцией пирувата. Окислительное декарбоксилирование пирувата до ацетил-КоА. СО2 осуществляется пируватдегидрогеназным комплексом и сопровождается накоплением в цитоплазме НАД • Н2. Последний используется для генерирования Н+ в процессе секреции НС1. Расщепление триглицеридов в слизистой оболочке желудка под влиянием триглицеридлипазы и последующая утилизация жирных кислот создает в 3—4 раза больший приток восстановительных эквивалентов в митохондриальную цепь переноса электронов. Как аэробный гликолиз, так и окисление жирных кислот запускаются посредством цАМФ-зависимого фосфорилирования соответствующих ферментов, обеспечивающих генерирование ацетил-КоА в цикле Кребса и восстановительных эквивалентов для электронпереносящей цепи митохондрий. Са2+ является необходимым элементом секреторной системы НС1.
Процесс цАМФ-зависимого фосфорилирования обеспечивает активацию желудочной карбоангидразы, которая является регулятором кислотноосновного равновесия в кислотопродуцирующих клетках. Работа этих клеток сопровождается длительной и массовой потерей ионов Н+, что приводит к накоплению в клетке ОН-, способных оказать повреждающее действие на клеточные структуры. Нейтрализация гидроксильных ионов и является главной функцией карбоангидразы. Образующиеся бикарбонатные ионы посредством электронейтрального механизма выводятся в кровь, а ионы СГ входят в клетку.
Кислотопродуцирующие клетки на наружных мембранах имеют две мембранные ферментные системы, участвующие в механизмах продукции Н+ и секреции НС1. Ими являются Ка+-К+-АТФаза и Н+-К+-АТФаза. Иа+-К+-АТФаза, расположенная в базолатеральных мембранах клеток, переносит К+ из крови в обмен на Na+, а Н+-К+-АТФаза, локализованная в секреторной мембране, транспортирует калий из первичного секрета в обмен на выводимые в желудочный сок ионы Н+. Процесс образования соляной кислоты кислотопродуцирующими клетками схематически представлен на рис. 11.11.
В период секреции митохондрии всей массой охватывают в виде муфты секреторные канальцы, и их мембраны сливаются, образуя митохондриально-секреторный комплекс, где ионы Н+ непосредственно акцептируются Н+-К+-АТФазой секреторной мембраны и транспортируются из клетки.
Таким образом, кислотообразующая функция обкладочных клеток осуществляется благодаря процессу фосфорилирования — дефосфорилирования, наличию митохондриальной окислительной цепи, транспортирующей ионы Н+ из матриксного пространства, а также активности Н+-К+-АТФазы секреторной мембраны, перекачивающей протоны из клетки за счет энергии АТФ.
Вода поступает в канальцы клетки путем осмоса. Конечный секрет, поступающий в канальцы, содержит НС1 в концентрации 155 ммоль/л, хлористый калий в концентрации 15 ммоль/л и очень малое количество хлористого натрия.
Роль соляной кислоты в пищеварении. В полости желудка хлористоводородная кислота: 1) стимулирует секреторную активность желез желудка; 2) способствует превращению пепсиногена в пепсин путем отщепления ингибирующего белкового комплекса; 3) создает оптимальную кислотность для действия протеолитических ферментов желудочного сока; 4) вызывает денатурацию и набухание белков (что способствует их расщеплению ферментами); 5) обеспечивает антибактериальный эффект секрета; 6) участвует в осуществлении механизма перехода пищи из желудка в двенадцатиперстную кишку, раздражая хеморецепторы ее слизистой оболочки; 7) участвует в регуляции секреции желудочных и поджелудочных желез, стимулируя образование гастроинтестинальных гормонов (гастрина, секретина); 8) возбуждает секрецию фермента энтерокиназы энтероцитами слизистой оболочки двенадцатиперстной кишки; 9) участвует в створаживании молока; 10) стимулирует моторную активность желудка.
Ферменты желудочного сока и их роль в пищеварении. В полости желудка под влиянием протеолитических ферментов осуществляется начальный гидролиз белков до альбумоз и пептонов. Протеолитические ферменты желудочного сока обладают активностью в широком диапазоне колебаний pH с оптимумом действия при pH 1,5—2,0 и 3,2—4,0. Это обеспечивает гидролиз белков в условиях значительных колебаний концентрации соляной кислоты в желудочном соке, в слоях пищи, прилежащих к слизистой оболочке желудка, и в глубине содержимого желудка.
В желудочном соке представлены семь видов пепсиногенов, объединенных общим названием пепсины. Образование пепсинов осуществляется из неактивных предшественников — пепсиногенов, находящихся в клетках желудочных желез в виде гранул зимогена. В просвете желудка пепсиноген активируется НС1 путем отщепления от него ингибирующего белкового комплекса. В дальнейшем в ходе секреции желудочного сока активация пепсиногена осуществляется аутокаталитически под действием уже образовавшегося пепсина.
При оптимальной величине pH среды пепсин осуществляет гидролиз белков, разрывая в белковой молекуле пептидные связи, образованные группами фениламина, тирозина, триптофана и других аминокислот. В результате этого белковая молекула распадается на пептоны и пептиды. Пепсин обеспечивает гидролиз основных белковых веществ, особенно коллагена — основного компонента волокон соединительной ткани,
К основным пепсинам желудочного сока относятся следующие.
Пепсин А — группа ферментов, гидролизирующих белки при оптимуме pH 1,5—2,0. Часть пепсиногена (около 1 %) переходит в кровеносное русло, откуда вследствие небольшого размера молекулы фермента проходит через клубочковый фильтр в почках и выделяется с мочой (уропепсиноген). Определение содержания уропепсина в моче используется в лабораторной практике для характеристики протеолитической активности желудочного сока.
Гастриксин (пепсин С), гидролизирующий белки при оптимуме pH 3,2— 3,5. Пепсин В (парапепсин) расщепляет желатину и белки соединительной ткани. При pH 5,6 и выше протеолитическое действие фермента ослабляется.
Реннин (пепсин Д, химозин) расщепляет казеин молока в присутствии ионов Са2+.
Желудочный сок содержит ряд непротеолитических ферментов. Среди них — желудочная липаза, расщепляющая жиры, которые находятся в пище в эмульгированном состоянии (жиры молока), на глицерин и жирные кислоты при pH 5,9—7,9. У грудных детей желудочная липаза расщепляет до 59 % жира молока. В желудочном соке взрослых людей липазы мало. Поэтому основное количество жиров переваривается в тонком кишечнике.
Клетками поверхностного эпителия слизистой оболочки желудка вырабатывается лизоцим (муромидаза). Лизоцим обусловливает бактерицидные свойства желудочного сока.
Уреаза расщепляет мочевину в желудке при pH 8,0. Освобождающийся при этом аммиак нейтрализует соляную кислоту и предотвращает избыточную кислотность химуса, поступающего из желудка в двенадцатиперстную кишку.
Желудочная слизь и ее значение. Органическим компонентом желудочного сока является слизь.
Нерастворимая слизь (муцин) является продуктом секреторной активности добавочных клеток и клеток поверхностного эпителия. Муцин выделяется через апикальную мембрану мукоцита, образует слой слизи толщиной 0,5—1,5 мм, он обволакивает слизистую оболочку желудка и препятствует повреждающему воздействию соляной кислоты и пепсинов на клетки слизистой оболочки и раздражающих веществ, поступивших с пищей. Этими же клетками одновременно с муцином продуцируется и бикарбонат. Образующийся при взаимодействии муцина и бикарбоната мукозо-бикар- бонатный барьер предохраняет слизистую оболочку от аутолиза под воздействием соляной кислоты и пепсинов.
Слой слизи является преградой для обратной диффизии ионов водорода из полости желудка; он нейтрализует соляную кислоту благодаря буферным свойствам из-за наличия гидрокарбонатов, а также адсорбирует ферменты. Под влиянием длительного воздействия желчных кислот (при забрасывании их из двенадцатиперстной кишки), салицилатов, масляной и пропионовой кислот, алкоголя происходит нарушение слизистого барьера. Это приводит к обратной диффизии ионов водорода из полости желудка, контакту слизистой оболочки с пепсинами и ее повреждению в результате аутолиза. Так формируются пептические язвы желудка. Возникновению язвенного процесса способствуют продукты жизнедеятельности микроорганизма Helicobacter pylori, которые усиливают секрецию соляной кислоты.
11.5.2. Регуляция секреции желудочного сока
Железы желудка в состоянии относительного покоя (в условиях отсутствия процесса пищеварения) выделяют небольшое количество сока нейтральной или слабощелочной реакции (фоновая секреция). Под влиянием пищевого раздражения во время приема пищи железы желудка секретируют значительный объем желудочного сока, богатого протеолитическими ферментами. Эта реакция желез является рефлекторным ответом на раздражение пищей рецепторов слизистой оболочки ротовой полости, глотки и желудка (безусловный рефлекс) и воздействие комплекса раздражителей, влияющих на другие рецепторы, предшествующих и сопутствующих приему пищи (условный рефлекс).
Афферентные импульсы с раздражаемых пищей рецепторов (тактильных, температурных и вкусовых) по афферентным волокнам V, VII, IX и X черепно-мозговых нервов передаются в бульбарный, таламический, гипоталамический и корковый отделы пищевого центра. Нисходящие влияния коркового представительства пищевого центра активируют парасимпатические и симпатические ядра гипоталамуса. При возбуждении парасимпатических ядер их эфферентные влияния активируют клетки бульбарного отдела пищевого центра, импульсы от которых по преганглионарным волокнам блуждающего нерва передаются на постганглионарные нейроны интрамуральных ганглиев желудка (клетки Догеля I типа). Выделяемый в окончаниях их аксонов медиатор ацетилхолин через М-холинорецепторы мембраны гландулоцитов стимулирует секреторную деятельность главных, обкладочных и мукоидных клеток желез желудка. Поэтому после перерезки блуждающих нервов у собак не выделяется желудочный сок при раздражении пищей рецепторов полости рта, на ее вид и запах.
Ацетилхолин активирует также эндокринные У-клетки слизистой оболочки антрального отдела желудка, вырабатывающие гастрин, который через кровоток достигает обкладочных клеток желез и через специальные рецепторы их мембраны вызывает выработку соляной кислоты.
Ацетилхолин и гастрин в слизистой оболочке фундального отдела желудка стимулируют активность эндокринных клеток, выделяющих гистамин, который, диффундируя по межклеточным пространствам, через Н2- рецепторы мембраны обкладочных клеток стимулирует выработку большого количества кислого желудочного сока, бедного ферментами и мукоидами. После перерезки блуждающих нервов (в экспериментах на животных) сокогонный эффект ацетилхолина, гастрина и гистамина резко ослабляется.
Поступление пищи в желудок во время еды усиливает безусловно-рефлекторное отделение желудочного сока, что обусловлено раздражением механорецепторов при растяжении его стенок (особенно антрального отдела) и хеморецепторов пептидами и экстрактивными веществами пищи. При этом рефлекс осуществляется с участием бульбарного отдела пищевого центра, дополняется местной реакцией, реализуемой через холинергические нейроны интраорганной нервной системы.
Увеличение концентрации соляной кислоты в желудочном соке приводит к угнетению выработки гастрина У-клетками и последующим уменьшением кислотности сока. Этот механизм саморегуляции предотвращает избыточное выделение соляной кислоты. Подавляют секрецию соляной кислоты соматостатин, ВИП и серотонин (см. табл. 11.1).
При возбуждении симпатических ядер гипоталамуса его нисходящие влияния стимулируют преганглионарные симпатические нейроны боковых рогов торакальных сегментов (IV—X) спинного мозга. Их эфферентные влияния стимулируют постганглионарные нейроны симпатических ганглиев, аксоны которых в составе чревных нервов достигают желез желудка. Выделяющийся в их окончаниях медиатор норадреналин тормозит секрецию желудочного сока, но приводит к накоплению секреторного материала в гландулоцитах, содержащих пепсиногены и мукоиды.
Поступление в двенадцатиперстную кишку не полностью переваренного химуса из желудка вызывает дополнительное возбуждающее и тормозное влияние на железы желудка. Гастрин, вырабатываемый У-клетками слизистой оболочки кишки, бомбезин (гастрин-рилизинг-гормон), выделяемый Р-клетками, продукты гидролиза белков гуморально стимулируют выработку соляной кислоты.
Выделяемые эндокринными клетками двенадцатиперстной кишки холецистокинин, гастроингибирующий пептид, нейротензин и другие пептиды (см. табл. 11.1) под влиянием продуктов гидролиза жиров в двенадцатиперстной кишке оказывают тормозной эффект на выработку соляной кислоты железами желудка.
11.5.2.1.Фазы желудочной секреции
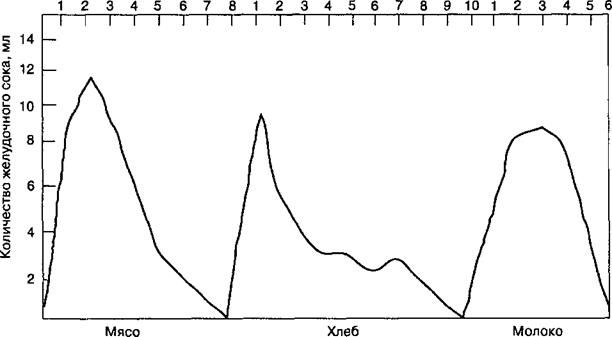
В естественных условиях приема пищи сокоотделение в желудке начинается под влиянием условных раздражителей — вида и запаха пищи, привычной обстановки. При попадании пищи в ротовую полость в процессе жевания и глотания раздражаются рецепторы слизистой оболочки рта, глотки и пищевода, что вызывает безусловно-рефлекторную секрецию желудочного сока. Этот комплекс условных и безусловных секреторных рефлексов составляет сложнорефлекторную первую фазу желудочного сокоотделения, которая продолжается около двух часов. Выделяющийся при этом сок отличается высоким содержанием ферментов, что способствует возникновению второй фазы сокоотделения — нейрогуморальной. Она обусловлена раздражением рецепторов желудка и кишечника компонентами химуса, влиянием на железы желудка гастроинтестинальных гормонов (см. табл.
11.1) и продуктов гидролиза пищевых веществ, поступающих в кровоток. Поэтому секреция желудочного сока (в опытах на собаках) продолжается 6—10 ч.
Нейрогуморальная фаза секреции желудочного сока «наслаивается» на сложнорефлекторную и состоит из желудочной и кишечной фаз. Желудочная фаза сокоотделения обусловлена влиянием содержимого желудка на рецепторы его слизистой оболочки и эндокринные клетки (см. табл. 11.1). Так, механическое раздражение пищей приводит к выделению гастрина. Его выделение усиливается под влиянием продуктов гидролиза белков и экстрактивных веществ.
Кишечная фаза секреции желудочного сока обусловлена раздражением рецепторов слизистой оболочки двенадцатиперстной кишки кислым желудочным содержимым, поступающим из желудка, всасыванием в кровь продуктов гидролиза белков и гастроинтестинальных гормонов, выделяемых эндокринными элементами слизистой оболочки кишечника (см. табл.
11.1). При осуществлении кишечной фазы возникают и тормозные влия-
Часы