 2020-05-25
2020-05-25 363
363Рецепторы могут реагировать на разные параметры раздражения, включение и выключение, напряжение и убывание силы раздражения и т.д.
Передаточная функция рецепторов. Установлено, что количественные соотношения между амплитудой стимула и частотой выходящих из рецепторов импульсаций (передаточная функция) не одинаковы в разных рецепторах. Медленный рецептор растяжения может иметь линейную передаточную функцию в широком диапазоне, т.е. частота потенциалов действия этих рецепторов пропорциональна амплитуде стимулов.
Многие другие рецепторы имеют передаточную функцию, при которой рецепторный потенциал (а при надпороговых стимулах и частота потенциалов действия) быстро возрастает при низкой амплитуде стимула, но чувствительность рецептора постепенно снижается с увеличением амплитуды стимула. Такие процессы наблюдаются, например, в фоторецепторах.
У болевых рецепторов частота потенциалов действия нарастающе увеличивается по мере увеличения интенсивности стимула.
Следует отметить, что информационная функция рецепторов в классической физиологии исследована с традиционных позиций действия стимулов. Причем очень часто эти эксперименты проводятся на изолированных переживающих препаратах рецепторов.
Системные свойства рецепторов. В целом организме свойства рецепторов, включенных в различные функциональные системы, качественно отличаются. Многочисленные рецепторы (особенно внешние), наряду с реакцией на внешние стимулы, активно настраиваются доминирующей мотивацией на специальные раздражители.
Кодирование информации рецепторами. В соответствии с принципом двоичного кодирования информации передача информации о действии стимула на рецептор определяется его интенсивностью. Если на выходе рецептора отмечается отсутствие импульса или один импульс, то его отсутствие несет информацию о том, что интенсивность раздражения ниже порога, тогда как наличие одного импульса означает, что интенсивность выше порога. При увеличении числа импульсов на выходе рецепторов можно судить о разных уровнях стимуляции. Если максимальное число импульсов
в афферентном волокне равно N, то теоретически рецептор может передать в ЦНС сигналы о N+ 1 различных уровнях интенсивности стимула.
Число различимых уровней интенсивности стимула, кодируемых сигналом, передаваемым по нервному волокну, для идеального рецептора, который отвечает на постоянно действующий стимул импульсацией с постоянной частотой, равно N + 1 = fmt + 1, где fm — максимальная частота разряда рецептора, t — время наблюдения. Верхний предел /т определяется временем рефрактерности нервного волокна.
Амплитудно-частотный принцип передачи информации по нерву проявляется также в сальтаторном механизме. Сальтаторный механизм передачи возбуждения по нерву включает свойства волны (распространение электромагнитных волн через несколько соседних перехватов Ранвье) и частиц — перемещений ионов вдоль мембран.
Для нервной системы типична избыточность передачи информации. Это — параллельная передача информации по двум или большему числу каналов. Кроме того, избыточность передачи информации определяется плотностью расположения рецепторов на периферии. Все это компенсирует помехи процессу кодирования в рецепторах. Механизм латерального торможения (см. гл. 3) также ограничивает ширину распространения сигналов при передаче их по параллельным каналам к ЦНС.
Функциональная мобильность рецепторов. В каждый определенный момент времени не все множество рецепторов одной модальности реагирует на внешнее раздражение. Например, из 250 тыс. чувствительных к холоду рецепторов в определенный момент реагирует только 50 тыс. При воздействии на организм света или тепла количество терморецепторов, чувствительных к теплу, возрастает. Способность рецепторов реагировать на раздражение не всей массой составляющих его элементов, а парциально, частями, и изменять реакции при определенных воздействиях называется функциональной мобильностью (П. Г. Синякин). Функциональная мобильность обеспечивает значительное снижение утомляемости органов чувств и резервирование рецепторов.
Существенная роль в повышении функциональной мобильности рецепторов принадлежит доминирующей мотивации.
Рецептивные поля. Определенное множество рецепторов, связанных с отдельным афферентным волокном, называется рецептивным полем.
Рецептивные поля первичночувствующих рецепторов организованы наиболее просто. Например, тактильное или ноцицептивное рецептивное поле кожной поверхности представляет собой разветвления одиночного чувствительного волокна.
Рецепторы, расположенные в различных участках рецептивного поля, имеют различную чувствительность к адекватному раздражению. В центре рецептивного поля обычно находится высокочувствительная зона, а ближе к периферии рецептивного поля чувствительность падает.
Рецептивные поля вторичночувствующихрецепторов организованы аналогичным образом. Отличие состоит в том, что разветвления афферентного волокна оканчиваются не свободно, а имеют синаптические контакты с чувствительными клетками — рецепторами. Так организованы вкусовые, вестибулярные, акустические рецептивные поля.
Более сложную организацию имеют рецептивные поля сетчатки глаза. Ганглиозные клетки, передающие информацию от рецепторов в ЦНС, связаны с фоторецепторами посредством так называемых биполярных клеток.
Свойства рецептивных полей. Если на отдельный рецептор (например, кожного механорецептивного поля) с определенным интервалом повторно наносить адекватное механическое раздражение, то ответ, регистрируемый с афферентного волокна, постепенно уменьшается и исчезает. Привыкание, или инактивация, может длиться от нескольких секунд (механорецепторы) до 3—5 мин (ноцицепторы). Если раздражающий стимул переместить в другую точку поля, то он снова вызывает ответ. Такая «дееспособность» других рецепторов рецептивного поля при выключении одного из них обеспечивает надежность функционирования рецептивного поля.
При одновременном раздражении двух различных рецепторов одного рецептивного поля с афферентного волокна можно получить лишь один ответ от рецептора, разряжающегося с более коротким латентным периодом или с большей частотой, чем второй. Даже при максимальном раздражении всех рецепторов поля на афферентное волокно всегда «выходит» ответ только одного рецептора.
Более сложные отношения между центром рецептивного поля и периферией имеют место в зрительных рецептивных полях — торможение рецепторов на периферии поля и активация рецепторов в центре, и наоборот.
Перекрытие рецептивных полей. Один и тот же участок чувствительной поверхности (например, кожи или сетчатки глаза) иннервируется несколькими чувствительными нервными волокнами, которые своими разветвлениями перекрывают рецептивные поля отдельных афферентных нервов. Особенно выражено такое перекрытие на дистальных и проксимальных частях кожи конечностей, т.е. на участках тела, наиболее подверженных действию механических раздражений и занимающих особое место в их анализе (ладонные поверхности у человека и обезьяны, подушечки лап и морда у кошки). Перекрытие рецептивных полей может быть 10—40-кратным, в результате чего относительно увеличивается общая сенсорная поверхность организма, возрастает дублирование каналов информации.
Благодаря перекрытию рецептивных полей осуществляется пространственное восприятие раздражения.
Периферическое взаимодействие рецепторов. Афферентный разряд импульсов, возникающий под действием адекватного стимула, не только распространяется по нервному волокну в ЦНС, но и по разветвлениям волокна может поступать к соседним, окружающим рецептивное поле рецепторам, вызывая их активацию или торможение, так называемый аксон-рефлекс.
1.8. ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
Обмен веществ и энергии — основное свойство живых существ. В процессе обмена питательные вещества превращаются в собственные компоненты тканей и конечные продукты метаболизма. Эти процессы сопровождаются поглощением и выделением энергии. Обмен веществ определяет важный физиологический процесс — формирование метаболических потребностей организма.
Обмен веществ. Обмен веществ в организме обычно подробно излагается в курсе биохимии. Здесь будут рассмотрены только некоторые важнейшие физиологические стороны обмена веществ.
Питательные вещества, поступающие в организм, несут пластическую и энергетическую нагрузку. Пластическую роль в орга
низме, связанную с построением тканей, играют в основном белки. Энергетическую роль играют углеводы, жиры и, в незначительной степени,белки.
Потребности в питательных веществах варьируют в зависимости от генетической конституции субъектов, размеров их тела, возраста, пола, активности желез внутренней секреции, физической активности, репродуктивной функции и т.д.
Для нормального обмена веществ кроме белков, жиров и углеводов абсолютно необходимы вода, минеральные соли, микроэлементы и витамины.
Средняя суточная потребность в белке — 80— 100 г, углеводах — 500 г, жирах — 70—80 г.
При всех видоизменениях поступающих в организм веществ в нормальном организме общее количество потребленных веществ равно выделяемому.
Обмен энергии. В основе процессов обмена энергии лежат законы термодинамики. С точки зрения термодинамики живые существа относятся к открытым стационарным неравновесным системам. Они обмениваются с окружающей средой веществом и энергией. С другой стороны, благодаря механизмам саморегуляции составляющих их функциональных систем живые организмы удерживают свои основные функции и энергетические затраты на них в определенных пределах.
Принцип устойчивого неравновесия живых систем (Э. С. Бауер) гласит: живые системы никогда не бывают в равновесии с окружающей средой и исполняют за счет своей внутренней энергии постоянную работу против равновесия, требуемого законами физики и химии.
Законы термодинамики
Первый закон термодинамики — закон сохранения и превращения энергии (М. В. Ломоносов, А. Лавуазье) гласит: энергия не исчезает и не образуется вновь, а только переходит из одной формы в другую — энергия механической работы, кинетическая энергия и теплота могут превращаться друг в друга.
Второй закон термодинамики (Больцман): часть энергии организма способна к превращениям и совершению полезной работы. Другая часть — рассеивается в виде тепла, характеризуя меру термодинамической неупорядоченности системы — энтропию.
Живые организмы не могут обратно использовать выделяющуюся в процессах жизнедеятельности тепловую энергию.
Второй закон термодинамики утверждает, что энтропия может только возрастать или оставаться постоянной.
Тем не менее живые организмы, благодаря саморегуляции, работают против энтропии. Они представляют так называемые «ква- зизакрытые» системы (И. Р. Пригожин, И. Стенгере).
Поступление энергии в организм. Свободная энергия поступает в организм в основном с питательными энергетическими веществами, а также с солнечной энергией. Энергия, заключенная в поступающих в организм веществах, трансформируется в клетках организма в химическую, электрическую, механическую и осмотическую энергию. Часть энергии выделяется из организма в виде тепла.
Энергия солнечных лучей стимулирует синтез биологически активных веществ в коже, сетчатке глаза, а также вызывает изменения в деятельности желез внутренней секреции.
Перенос энергии в клетках. Важнейший переносчик энергии в клетках — аденозинтрифосфорная кислота (АТФ). Потенциальная энергия, содержащаяся, например, в химических связях молекул глюкозы, переходит в результате аэробного или анаэробного окисления в форму богатой энергией фосфатной связи АТФ.
Солнце — источник внеатомной электромагнитной энергии. При столкновении фотонов (квантов света) с электронами, находящимися на какой-либо орбите, за счет приобретения электронами дополнительной энергии этих квантов происходит увеличение внутренней энергии атомов и переход их в возбужденное состояние. Коротковолновая часть электромагнитного спектра, излучаемого Солнцем, которая включает рентгеновские лучи, у-лучи, ультрафиолетовые лучи, обладает высокой энергией и способна повреждать живую ткань. Видимый свет, а также инфракрасное излучение обладают относительно низкой энергией, но способны переводить электроны, которые находятся на внешних орбитах, на более высокие энергетические уровни. Переходя на низшую орбиту, электроны делают атом возбужденным, выделяют энергию и могут вступать в различные химические реакции.
Полагают, что в основе энергетических процессов жизнедеятельности лежат сложные перемещения электронов (А. Сент-Дьерди).
Энергозатраты у человека зависят от пола, возраста, массы, интенсивности выполняемой работы, от времени суток, сезонов и др. Они меняются в связи с изменением окружающей температуры, влажности, барометрического давления и пр.
Поступающая в организм энергия используется в процессах клеточного метаболизма, мышечной деятельности, секреторной деятельности и в процессах нервного возбуждения.
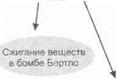
Калорическая ценность питательных веществ. Калорическую ценность питательных веществ определяют путем сжигания ] г вещества в специальном калориметре (бомба Бертло) при пропускании электрического тока. Калориметр погружается в воду, и измеряется разность температур воды до и после сжигания веществ. Установлено, что калорическая ценность 1 г белка равна 5,6 ккал, 1 г жира — 9,3 ккал, 1 г углеводов — 4,1 ккал.
Жиры и углеводы сгорают в бомбе Бертло и в организме до воды и углекислого газа. Белки в бомбе Бертло сгорают до конечных продуктов — N2, СО2 и Н2О. Однако в организме белки окисляются до мочевины, креатинина и мочевой кислоты. 1 г мочевины при сгорании выделяет 1,3 ккал. Поэтому физиологическая ценность белка в организме равна 4,1 ккал.
Указанные расчеты относительны. Трудно определить, сколько питательного продукта усваивается в каждом случае в организме. К тому же определенное количество принимаемых питательных веществ аккумулируется в депонированном состоянии.
Для расчета энергетической ценности принимаемых веществ необходимо количество белков, жиров и углеводов помножить на их физиологическую ценность.
Химическая калориметрия. Основана на окислении пищевых веществ сильными окислителями с известным количеством кислорода. После реакции избыток кислорода отгитровывается йодометрически.
Правило изодинамии Рубнера. Исходя из физических энергетических эквивалентов пищевых веществ, немецкий физиолог Рубнер усмотрел возможность их замены в соответствии с содержанием в них килокалорий тепла. Однако это правило не универсально. Жиры в определенной степени возможно заменять углеводами в соответствии с их калорической значимостью. Углеводы нельзя заменять жирами — «жиры сгорают в пламени углеводов». Белки
играют пластическую роль в организме, поэтому их также нельзя заменять другими веществами.
Калорические эквиваленты 1 л кислорода при окислении в организме различных веществ. Установлено, что при окислении в организме углеводов калорический эквивалент 1 л кислорода равен 5 ккал, при окислении жиров — 4,6 ккал, при окислении белков — 4,85 ккал.
Расчетным путем составлены таблицы зависимости между величинами дыхательных коэффициентов при окислении только жиров и углеводов и калорическим эквивалентом 1 л кислорода.
Определение энергозатрат организма. Когда не совершается физическая работа, вся химическая энергия переходит в тепловую. Это позволяет по количеству выделяемого организмом тепла судить об интенсивности энергетического обмена.
Количество тепла, выделяемого организмом, рассчитывают методом прямой и непрямой калориметрии.
Прямая калориметрия. Основана на непосредственном и полном учете количества выделенного организмом тепла с помощью специальных камер — биокалориметров. Первый калориметр для измерения энергозатрат у животных был изобретен в 1893 г. В. В. Пашутиным и А. А. Лихачевым.
Калориметры герметизированы и надежно теплоизолированы от окружающей среды. Для расчета количества тепла, выделяемого помещенным в калориметр животным или человеком, измеряют изменения температуры жидкости, протекающей через камеру за единицу времени.
Современные биокалориметры представляют собой специальные костюмы, снабженные термочувствительными датчиками.
Непрямая калориметрия. Газовый анализ. При полном газовом анализе энергозатраты рассчитывают за единицу времени по количеству поглощенного испытуемым кислорода и выделенной двуокиси углерода.
Отношение объема выделенной двуокиси углерода к объему поглощенного кислорода называется дыхательным коэффициентом.
Дыхательный коэффициент позволяет определить, за счет каких веществ преимущественно идет обмен веществ у испытуемых.
При дыхательном коэффициенте, равном 1, обмен веществ преимущественно осуществляется за счет окисления углеводов:
С6Н|2О(, + 60, = 6СО, + 6Н,О.
При дыхательном коэффициенте ниже 0,7 обмен веществ осуществляется преимущественно за счет окисления жиров:
2C5IH,SO6 + 1450, = Ю2СО, + 98Н,О.
Определенную трудность представляют расчеты энергозатрат при дыхательном коэффициенте 0,85, поскольку это могут быть энергозатраты при окислении белков или при окислении смеси жиров и углеводов. Это особенно важно для того, чтобы узнать, какую величину калорийной ценности кислорода при этом использовать — 4,85 ккал для белка или 4,85 для жиров и углеводов. Для исключения этих ошибок определяют «чистый» дыхательный коэффициент.
Для определения «чистого» дыхательного коэффициента необходимо исключить из окислительных процессов белок. Для этого определяют количество выделенного из организма с мочой азота. В белке — 16 % азота, т.е. 1 г азота содержится в 6,25 г белка. При умножении количества азота, выделенного за сутки, на 6,25 г получают количество белка, участвовавшего в суточных энергозатратах.
1 г белка при сгорании связывает 0,95 л О, и выделяет 0,76 л СО,. При умножении этих показателей на количество белка, участвовавшего в суточных энергозатратах, определяют, сколько О, пошло в сутки на окисление белков и сколько при этом выделилось СО,. Эти цифры вычитают, соответственно, из общего количества О, и СО,, в результате чего оказывается возможным определить «чистый» дыхательный коэффициент как отношение оставшегося СО, к О,.
Пример расчетов на основе полного газового анализа суточного расхода энергии у испытуемых
Исходные данные. За сутки испытуемый поглотил О, — 600 л, выделил СО, — 480 л. Азот мочи составил 16 г.
1. Поскольку 1 г азота мочи содержится в 6,25 г белка, за сутки окислилось 16x6,25 = 100 г белка.
2. 1 г белка при сгорании связывает 0,95 л О, и выделяет 0,76 л СО,. Отсюда на окисление 100 г белка пошло:
0,95 л х 100 = 95 л О,.
При этом выделилось:
' 0,76 л х 100 = 76 л СО,.
3. На окисление жиров и углеводов пошло кислорода:
600 л - 95 л = 505 л.
4. При окислении углеводов и жиров выделилось СО,:
480 л - 76 л = 404 л.
5. При этом «чистый» дыхательный коэффициент равен:
О, 505 л
Установленному дыхательному коэффициенту соответствует калорийная ценность 1 л О, 4,8 ккал.
6. При окислении углеводов и жиров выделилось:
4,8 ккал х 505 = 2424 ккал.
7. При окислении белка выделилось:
4,1 ккал х 100 = 410 ккал.
8. Общие энергозатраты за сутки:
2424 ккал + 410 ккал = 2834 ккал.
В данном случае энергозатраты осуществлялись у испытуемого преимущественно за счет окисления жиров и углеводов.
Неполный газовый анализ. При неполном газовом анализе расход энергии определяют по количеству кислорода, потребленного испытуемым за единицу времени. Для этой цели используют спирометаболографы, поглощающие в выдыхаемом воздухе СО, и позволяющие определять количество поглощенного испытуемыми О2.
Зная объем поглощенного за 1 мин кислорода (Vo,), усредненный дыхательный коэффициент при употреблении обычно смешанной пищи (0,85) и соответствующий ему калорический эквивалент кислорода (КЭК), равный 4,8 ккал, энергообмен рассчитывается по следующей формуле:
Z = Vo2 х КЭК = Vo2 х 4,8,
где КЭК — калорический эквивалент 1 л О2.
Как правило, неполный газовый анализ применяют для определения энергозатрат при основном обмене. В этом случае при исключении из энергообмена белков дыхательный коэффициент соответствует 0,85, т.е. только окисленной смеси углеводов и жиров.
На рис. 1.24 в схематической форме представлены различные виды калориметрии.
Основной обмен. Основной обмен — энергозатраты, связанные с процессами клеточного метаболизма.
В основной обмен вынужденно включаются энергозатраты жизненно важных процессов: деятельности сердца, дыхания, кровообращения, выделения, деятельности печени, почек, желез вну-
КАЛОРИМЕТРИЯ
 | |||||||
 | |||||||
 | |||||||
 | |||||||
| ||||
| ||||
 | ||||||||||||
 | ||||||||||||
 | ||||||||||||
 | ||||||||||||
 | ||||||||||||
 | ||||||||||||
 | ||||||||||||
(Определение
дыхательного
коэффициента)
Учитывается
количество
окислительного
белка
Определение
его чистого
дыхательного
коэффициента
Рис. 1.24. Различные виды калориметрии








