 2020-05-25
2020-05-25 1995
19951- й закон. Пороговое раздражение зависит не только от абсолютной величины раздражителя, но и от крутизны нарастания раздражения во времени.
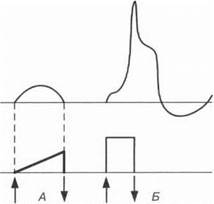 Закон сформулирован французским физиологом Э. Дюбуа-Рей- моном, который обнаружил, что ткань отвечает на раздражение при
Закон сформулирован французским физиологом Э. Дюбуа-Рей- моном, который обнаружил, что ткань отвечает на раздражение при
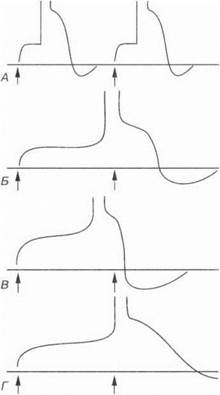
Рис. 1.2. При медленном нарастании тока в ткани регистрируется только местный потенциал (Д). При быстром нарастании напряжения раздражающего тока возникает потенциал действия (Б). Стрелки — включение и выключение электрического тока
резком нарастании тока пороговой силы. Ток пороговой силы при медленном нарастании не оказывает раздражающего действия на ткань. Крутизна фронта раздражающего тока является, таким образом, критической для раздражения тканей (рис. 1.2).
2- й закон. Раздражение зависит от времени действия раздражителя.
Установлено, что пороговое раздражение, произведенное кратковременно, становится подпороговым. Следовательно, для ответной реакции ткани важное значение имеет время раздражения.
Зависимость между величиной раздражения и временем его действия отражает кривая «силы-времени» (рис. 1.3).

Рис. 1.3. Кривая Вейса—Лапика «силы-времени»: по оси абсцисс — время действия раздражителя; по оси ординат — величина раздражения (напряжение тока)
Кривая «силы-времени» постулирует:
• для того чтобы вызвать ответную реакцию ткани, пороговый раздражитель должен иметь определенное время действия;
• если время действия порогового раздражителя меньше критического, раздражитель становится подпороговым;
• для надпороговых раздражителей для ответной реакции ткани необходимо меньшее время действия.
Пороговая, минимальная величина напряжения тока, способная вызвать возбуждение ткани, получила название реобаза.
Время, в течение которого электрический ток величиной в одну реобазу вызывает ответ ткани, называется полезным временем.
Измерение возбудимости ткани по порогу не точно. Пороговая величина раздражителя все время изменяется в соответствии с изменением функционального состояния ткани. При незначительном изменении порога время его действия меняется значительно. Поэтому предложено измерять возбудимость тканей другой условной величиной — хронаксией.
Хронаксия — время, в течение которого ток, равный удвоенной реобазе, вызывает ответную реакцию ткани. При удвоенной реобазе точка раздражения приходится на крутую часть кривой «силы-времени», в результате при незначительных изменениях возбудимости тканей она значительно не изменяется (см. рис. 1.3).
Кривые «силы-времени» и хронаксия различны у различных тканей и разных живых существ. У человека, например, хронаксия измеряется в миллисекундах. У гладкой мышцы улиток и желудка лягушек — в десятых и сотых долях секунды.
Измерение хронаксии получило название хронаксиметрии.
В практическом плане хронаксия имеет ряд недостатков:
• в целой мышце и нерве, используя хронаксию, нельзя определить поражение отдельных мышечных и нервных волокон;
• хронаксия характеризует только реакцию возбудимой ткани на одиночный стимул и не параметризует реакцию ткани на ритмический раздражитель.
Функциональная лабильность — способность возбудимых тканей воспроизводить частоту поступающих на них раздражений без трансформации (Н. Е. Введенский). Различные ткани обладают разной функциональной лабильностью. Наиболее лабильна нервная ткань. Так, диафрагмальный нерв может воспроизводить частоту импульсов без трансформации до тысячи в секунду.
Функциональная лабильность соматических нервов — 200— 300 имп./с, поперечно-полосатых мышц — 100—200 имп./с. Наиболее низкая функциональная лабильность у нервно-мышечных синапсов — 30—50 имп./с.
Оптимум и пессимум раздражения. Если ритм раздражений соответствует функциональной лабильности возбудимой ткани, то
ткань отвечает на каждый стимул. Состояние возбудимой ткани при этом получило название оптимума. В случае, когда частота подаваемых раздражений превышает функциональную лабильность ткани, в ней развивается состояние пессимума (рис. 1.4). Оптимум и пес- симум могут развиваться также при уменьшении или увеличении силы раздражения.
Пессимум
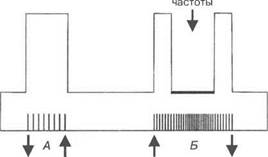
Рис. 1.4. Оптимум и пессимум сокращения мышцы:
А — оптимум сокращения мышцы при раздражении, частота которого соответствует ее функциональной лабильности; Б— при частоте раздражения, не соответствующей функциональной лабильности мышцы, она перестает отвечать на раздражение, развивается состояние пессимума. Стрелки — включение и выключение раздражения. Вертикальные черточки — частота раздражающих импульсов
Механизм оптимума и пессимума. Механизм пессимума обусловлен продолжительностью абсолютной рефрактерной фазы возбудимой ткани. Механизм оптимума определяется продолжительностью фазы супернормальной возбудимости ткани (экзальтации):
• если последующие раздражающие импульсы попадают в период абсолютной рефрактерности от предыдущих, новый цикл возбуждения не возникает;
• если последующие раздражающие импульсы попадают в период супернормальной возбудимости от предшествующих циклов возбуждения, ответная реакция возрастает до оптимальной величины.
3- й закон. Раздражение зависит от продолжительности действия раздражителя во времени.
Изменение порога раздражения во времени получило название аккомодация. Это еще один фактор, характеризующий ответ возбудимой ткани на раздражение. Любая ткань реагирует на внешнее воздействие постепенным повышением порога возбудимости.
Аккомодация определяет повышение порога раздражения в зависимости от скорости (крутизны) нарастания силы раздражителя. При медленном нарастании тока он может не вызвать возбуждения вследствие снижения возбудимости ткани (рис. 1.5). В основе явления аккомодации лежит инактивация натриевой и повышение калиевой проводимости мембраны возбудимой ткани.
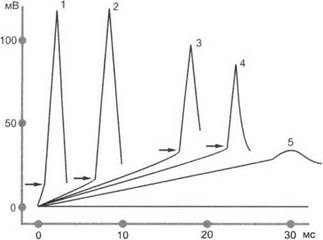
Рис. 1.5. Изменение амплитуды потенциалов действия изолированного нервного волокна при нарастании раздражающего тока: по оси абсцисс — время; по оси ординат — напряжение раздражающего тока
Разные ткани обладают разной аккомодацией. Чувствительные нервы обладают меньшей аккомодацией по сравнению с двигательными. Болевые нервные волокна не обладают аккомодацией.
Особенно отчетливо аккомодация проявляется при действии на ткань постоянного тока. При действии постоянного тока ответ ткани наблюдается только при замыкании и размыкании цепи тока.
Раздражение тканей постоянным током имеет следующие особенности.
Полярный закон (Э. Пфлюгер). Закон устанавливает место возникновения возбуждения в возбудимых тканях при действии постоянного тока:
• при замыкании цепи постоянного тока возбуждение возникает под катодом;
• при размыкании цепи постоянного тока возбуждение возникает на аноде.
Закон физиологического электротона. При начальном прохождении постоянного тока через возбудимую ткань под катодом возбудимость повышается (катэлектротон), а под анодом — понижается (анэлектротон). При продолжающемся действии постоянного тока под катодом развивается понижение возбудимости (ка- тодическая депрессия), а под анодом — повышение возбудимости (анодная экзальтация).
Закон «Все или ничего» — способность возбудимой ткани отвечать максимальной реакцией на пороговые и сверхпороговые раздражители.
При пороговых и сверхпороговых раздражениях возбудимые ткани порождают потенциал действия, который имеет постоянную величину. Ткань выдает максимум своих возможностей. При подпороговых воздействиях местный потенциал имеет градуальный характер.
Парабиоз — устойчивая длительная деполяризация мембраны, полное снижение ее возбудимости (Н. Е. Введенский). Факторы, вызывающие парабиоз (нейротоксины, наркотики, химические агенты, нагревание, охлаждение и пр.), называются альтерирующими, они полностью блокируют ионные каналы возбудимых мембран. Парабиоз, возникающий при действии на возбудимые ткани (например, на нерв альтерирующих факторов), представляет особую форму «застойного» нераспространяющегося возбуждения. После устранения действия альтерирующих факторов ткань восстанавливает свои исходные свойства возбудимости и проводимости.
Н. Е. Введенский при ритмическом электрическом раздражении нерва (на нервно-мышечном препарате при наложении на нерв хлористого калия) обнаружил три фазы в развитии парабиоза.
Фазы парабиоза
1- я фаза — провизорная, или уравнительная. В эту фазу сильное раздражение вызывает слабые ответные реакции ткани, а слабые,
подпороговые раздражители по-прежнему вызывают характерные для них слабые ответы. Величина ответов на раздражители разной силы в эту стадию становится одинаковой.
2- я фаза — парадоксальная. В эту фазу сильные раздражители вызывают меньшие по величине ответы, в то время как слабые раздражители, наоборот, вызывают увеличенные по амплитуде ответы.
3- я фаза — тормозная. Характеризуется полной утратой проводимости нерва и полным отсутствием реакции на раздражение (рис. 1.6).
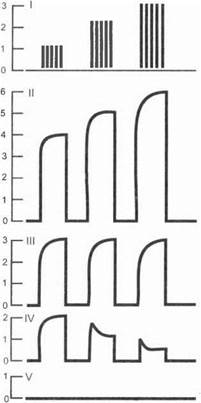 Рис. 1.6. Фазы парабиоза:
Рис. 1.6. Фазы парабиоза:
I — серии раздражений разной силы (слабые, средние, сильные);
II— V— ответные реакции на раздражения (// — до парабиоза,
III— уравнительная фаза, IV— парадоксальная фаза, V— тормозная
фаза)
В настоящее время в понятие парабиоза включаются все стадии.
Электрографическая характеристика парабиоза. На рис. 1.7 показана динамика изменения одиночного цикла возбуждения в участке альтерации по мере развития парабиоза.
Рис. 1.7. Динамика изменений одиночного цикла возбуждения в участке альтерации по мере развития парабиоза:
А — равноценные ответы на два последующих раздражения; Б — второе раздражение попадает в фазу относительной рефрактерное™ от первого (уравнительная фаза); В— второе раздражение попадает в фазу экзальтации от первого (парадоксальная фаза); Г— второе раздражение попадает в фазу абсолютной рефрактерное™ от первого (тормозная фаза парабиоза). Стрелки — одиночные раздражения
 Как видно из рис. 1.7, по мере развития парабиотического состояния волна потенциала действия начинает растягиваться во времени за счет удлинения следовых явлений. При этом также увеличивается продолжительность абсолютной рефрактерной фазы. Если повторное раздражение попадает в период относительной рефрактерности, проявляется 1-я стадия парабиоза. По мере «растягивания» потенциала действия повторные импульсы попадают в фазу повышенной возбудимости от предшествующих раздражений. В этом случае наблюдается 2-я стадия парабиоза. Наконец, если повторные раздражения попадают в растянутую абсолютную рефрактерную фазу,
Как видно из рис. 1.7, по мере развития парабиотического состояния волна потенциала действия начинает растягиваться во времени за счет удлинения следовых явлений. При этом также увеличивается продолжительность абсолютной рефрактерной фазы. Если повторное раздражение попадает в период относительной рефрактерности, проявляется 1-я стадия парабиоза. По мере «растягивания» потенциала действия повторные импульсы попадают в фазу повышенной возбудимости от предшествующих раздражений. В этом случае наблюдается 2-я стадия парабиоза. Наконец, если повторные раздражения попадают в растянутую абсолютную рефрактерную фазу,
наблюдается 3-я стадия парабиоза. Таким образом, парабиоз представляет «застойную» волну возбуждения. Эти «застойные» волны возбуждения не пропускают через место альтерации другие волны возбуждения. При этом в участке парабиоза замедлен ресинтез ацетилхолина, а также снижается уровень АТФ. Внешне при этом наблюдается торможение ответной реакции возбудимой ткани.
Н. Е. Введенский считал, что возбуждение и торможение по своей природе едины.
1.3. ВОЗБУЖДЕНИЕ
Возбуждение — процесс, развивающийся в возбудимой ткани под действием раздражителя.
Различают местный и распространяющийся процесс возбуждения. Местный процесс развивается в возбудимой ткани при действии подпороговых раздражителей. При достаточной силе раздражения процесс местного возбуждения переходит во взрывной процесс — распространяющийся процесс возбуждения.
Возбуждение возбудимых тканей характеризуется:
• изменением ионной проницаемости их мембран;
• электрическими процессами;
• работой метаболических насосов;
• изменением температуры возбужденного участка;
• изменением обмена веществ;
• изменением возбудимости.
Процесс возбуждения наиболее удовлетворительно объясняет мембранно-ионная теория (А. Ходжкин, Э. Хаксли, Б. Катц). Согласно этой теории, все клетки организма окружены полупроницаемыми мембранами, обладающими специфической избирательной ионной проницаемостью.
Морфофункциональная организация ионселективных каналов мембраны. Установлено, что полупроницаемость мембран определяет наличие в них электрически потенциалзависимых (для ионов Na+, К+, Са2+ и СГ) и потенциалнезависимых химически управляемых (например, для ионов К+) каналов и ионных насосов.
Ионные каналы состоят из поры, воротного механизма, сенсора (индикатора) и селективного фильтра (рис. 1.8). Их состояние определяет напряжение ионов в самой мембране.
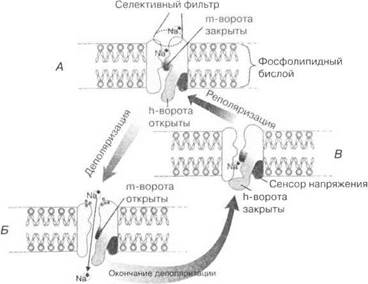
Рис. 1.8. Структура и механизм работы ионселективных каналов:
А — статическая поляризация, канал закрыт; Б — деполяризация, канал акти-
вирован; В — деполяризация, канал инактивирован
Пора— молекулярное динамическое образование, которое может находиться в открытом и закрытом состоянии. Поры образованы транспортными ферментами — белками с высокой каталитической активностью. Эти белки способны переносить ионы через мембраны со скоростью, в 200 раз превышающей скорость простой диффузии.
Воротный механизм — представлен двумя типами белковых молекул, расположенных на внешней (m-ворота) и на внутренней (h-ворота) сторонах мембраны. Благодаря свойству конформации — изменению пространственной конфигурации молекул без изменения их химической структуры — воротный механизм в тысячные доли секунды открывает/активирует (m-ворота) и закрывает (h-ворота) ионные каналы. Тем самым регулируется скорость продвижения ионов через мембранные каналы и обеспечивается квантовый характер поступления ионов в цитоплазму.
Сенсор напряжения ионов в мембране представлен белковой молекулой в самой мембране, способной реагировать на величину тока, протекающего в мембране.
Селективный фильтр располагается в наружной части ионных каналов. Он образован кольцом из отрицательно заряженных атомов кислорода. Эти заряды не пропускают анионы и притягивают катионы. Они определяют однонаправленное движение ионов через каналы и их избирательную проницаемость.
Одиночный цикл возбуждения
Процесс возбуждения — дискретный и распространяющийся. При действии раздражителя процесс возбуждения включает процессы деполяризации, реполяризации и гиперполяризации мембран и осуществляется импульсами, каждый из которых составляет одиночный цикл возбуждения. Одиночные циклы возбуждения являются информационными единицами. Из них складывается определенный рисунок, сообщение (паттерн) динамически распространяющегося по возбудимым тканям процесса возбуждения.
Электрическая характеристика одиночного цикла возбуждения. Одиночный цикл возбуждения в электрическом выражении состоит из следующих фаз (рис. 1.9): а) скрытый период; б) пред- спайк — местный, нераспространяющийся потенциал; в) спайк — распространяющийся потенциал (ток действия); г) отрицательный следовой потенциал; д) положительный следовой потенциал.
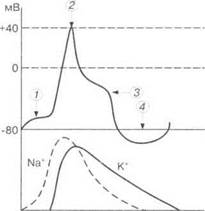 Рис. 1.9. Ионные токи Na+, К+
Рис. 1.9. Ионные токи Na+, К+
в соответствии с одиночным цик-
лом возбуждения:
1 — местный потенциал (предспайк);
2— спайк; 3— отрицательный сле-
довой потенциал; 4 — положитель-
ный следовой потенциал
Весь процесс одиночного цикла возбуждения составляет несколько миллисекунд (например, у нерва — от 1 до 10 мс). Ионная проницаемость мембраны определяет ее различные состояния: статическую поляризацию, деполяризацию, реполяризацию и гиперполяризацию.
Статическая поляризация — исходное состояние мембраны, предшествующее процессу возбуждения, состояние покоя. В состоянии покоя между наружной и внутренней поверхностью клеточных мембран существует разность потенциалов — мембранный потенциал, или потенциал покоя (калиевый потенциал).
Потенциал покоя, или мембранный потенциал, можно измерить, если один из отводящих электродов поместить на поверхности мембраны, а другой — ввести внутрь клетки. Разница напряжения между наружной и внутренней поверхностью мембраны — потенциал покоя — составляет 50—60 мВ.
Потенциал покоя определяется неравномерным распределением ионов между наружной и внутренней поверхностью мембраны.
Согласно мембранно-ионной теории, мембранный потенциал обусловлен неодинаковой проницаемостью мембраны возбудимых клеток для ионов Na+, К+ и СГ.
Цитоплазма нервных и мышечных клеток содержит в 30—50 раз больше ионов К+, в 8—15 раз меньше ионов Na+ и в 10—50 раз меньше ионов СГ по сравнению с внеклеточной жидкостью.
Неравномерное распределение ионов на мембранах в состоянии покоя определяется:
• наличием трансмембранных градиентов концентраций ионов Na+, К+ и СГ;
• высокой избирательной проницаемостью мембраны для ионов К+ благодаря тому, что в состоянии покоя постоянно открыты калиевые каналы;
• практически полной непроницаемостью мембраны для ионов Na+, для которых ионные каналы полностью закрыты (закрыты ш-ворота);
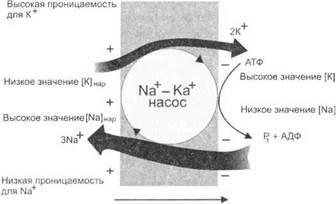
• работой Ка+,К?-АТФазных насосов, активно поддерживающих постоянные ионные градиенты концентрации, «выкачиванием» ионов Na+ наружу мембраны и обратным поступлением ионов К+ в цитоплазму (рис. 1.10). Ионный насос перемещает ионы против их концентрационного градиента и работает с затратой энергии АТФ.
Благодаря отмеченным механизмам состояние статической поляризации мембраны характеризуется избытком ионов Na+ на поверхности мембраны и ионов К+ внутри клетки (рис. 1.11).
При этом ионы СГ играют уравновешивающую роль, роль своеобразного «балансера». Как только изменяется натриево-калиевое равновесие, ионы СГ активно перемещаются через мембрану, исправляя нарушенный градиент ионов Na+ и К+ между наружной
Внеклеточная Внутриклеточная
среда среда
 | |||
 | |||
|

К; Na;5 Clw
к;0 Na; ci;
Рис. 1.11. Распределение ионов между внутренней и наружной
стороной мембраны возбудимой клетки
и внутренней поверхностью мембраны, активно поддерживаемый Na.K -АТФазным насосом.
Таким образом, состояние статической поляризации мембраны — активный физико-химический процесс, работающий против пассивного распределения ионов между полупроницаемыми мембранами по градиенту их концентрации.
В результате неравномерного распределения ионов между наружной и внутренней поверхностью мембраны ее наружная поверхность заряжена положительно по отношению к внутренней.
Деполяризация развивается в возбудимых тканях под действием раздражителей.
Раздражитель при действии на мембрану нарушает ее проницаемость, в результате чего изменяется присущее состоянию покоя ионное равновесие. Под действием раздражителя открываются поворота быстрых натриевых каналов (см. рис. 1.8, Б), вследствие чего ионы Na+ лавинообразно пассивно (по градиенту концентрации) поступают в клетку. В результате снижается величина положительного заряда на поверхности мембраны и увеличивается в цитоплазме. Указанные процессы лежат в основе формирования местных (локальных) потенциалов. Как только под продолжающимся действием раздражителя значение местного потенциала достигает критической точки и падает до нуля, происходит перезарядка мембраны и инверсия ее заряда (овершут). Поверхность мембраны становится электроотрицательной по отношению к цитоплазме. Происходит взрывной процесс — возникает потенциал действия — спайк.
Одновременно через открытые калиевые каналы начинают выходить ионы К+, возвращая мембранный потенциал к исходному уровню (см. рис. 1.9).
Спайк, или потенциал действия, — типичный триггерный механизм. Его происхождение определяется внутренней молекулярной организацией мембраны. В то же время электрический потенциал действия — причина деполяризации соседних пунктов мембраны, т.е. распространения возбуждений по возбудимым тканям.
Напряжение потенциала действия — около 120 мВ.
Факт внезапности (триггерный механизм) исключительно важен для последующего распространения процесса возбуждения по возбудимой ткани. Потенциал действия часто обозначают как распространяющееся возбуждение.
Деполяризация может быть вызванной действием на мембрану внешнего стимула и спонтанной у некоторых видов клеток-пейсме- керов, которые способны самовозбуждаться.
При достижении критического значения тока (120 мВ) сенсор захлопывает h-ворота натриевых каналов, прекращая тем самым дальнейшее поступление ионов Na+ в цитоплазму.
Ионные перемещения в мембране в процессе возбуждения впервые зарегистрированы на гигантском аксоне кальмара, из которого была выдавлена внутренняя эндоплазма.
Реполяризация — восстановление исходной статической поляризации мембраны. Процесс реполяризации мембраны характеризуется:
• активацией Ма+,К+-АТФазного насоса и активным транспортом ионов Na+ из клетки в межклеточную жидкость;
• пассивным транспортом ионов К+ в клетку по градиенту концентрации.
Реполяризация приводит к открыванию h-ворот и закрыванию m-ворот (см. рис. 1.8, Б).
Электрографическим проявлением реполяризации является нисходящая часть спайка и отрицательный следовой потенциал.
Гйперполяризация — увеличение уровня мембранного потенциала по сравнению с состоянием статической поляризации.
Гиперполяризация кратковременно наблюдается в процессе реполяризации вслед за восстановлением исходного мембранного потенциала. Гиперполяризация обусловлена повышением проницаемости калиевых каналов и каналов для ионов СГ. В связи с этим поверхность мембраны приобретает избыточный, по сравнению с состоянием покоя, положительный заряд. Электрографическим проявлением гиперполяризации является следовой положительный потенциал.
Метаболические процессы при возбуждении. Под действием ионных перемещений из прекурсорной, связанной с белком формы, в клетке образуется активный ацетилхолин, который также способствует процессу возбуждения. Процесс возбуждения обеспечивается энергией АТФ и креатинфосфата (рис. 1.12).
Потенциал действия регулирует выделение медиаторов в синапсах, внутриклеточную концентрацию кальция и сокращение мышечной ткани, выделение гормонов железами внутренней секреции и др.
Поглощение кислорода при возбуждении увеличивается по сравнению с состоянием покоя в среднем на 24%. Значительно возрастает теплопродукция: активируются ферменты клетки, участвующие в процессах аэробного и анаэробного обмена и окислительного фосфорилирования. В процессе возбуждения в результате
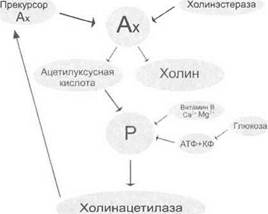
Рис. 1.12. Схема метаболических процессов, сопровождающих процесс возбуждения в возбудимых тканях. Ацетилхолин (Ах) под влиянием холинэстеразы расщепляется на ацетилуксусную кислоту и холин. Под влиянием АТФ, креатинфосфата, витамина В, ионов Са2* и Мд2+образуется фермент холинацетилаза, которая восстанавливает прекурсор Ах
перекисного окисления липидов мембраны образуются свободные радикалы.

