 2020-05-25
2020-05-25 2229
2229Местное возбуждение:
• возникает при действии подпороговых стимулов;
• возникает локально — только в зоне действия стимула;
• быстро «затухает» во времени;
• при повторных воздействиях способно к суммации и может перейти в распространяющееся возбуждение;
• характеризуется некоторым повышением возбудимости;
• электрографически характеризуется местным потенциалом (предспайком);
• не подчиняется закону «все или ничего».
Распространяющееся возбуждение:
• возникает при действии пороговых и надпороговых стимулов или в результате суммации до критического уровня местных возбуждений;
• распространяется на значительные расстояния от места возникновения;
• не «затухает» во времени — бездекрементное распространение;
• не способно к суммации;
• сопровождается циклическими изменениями возбудимости с наличием абсолютной рефрактерной фазы;
• электрографически характеризуется потенциалом действия (спайком);
• подчиняется закону «Все или ничего».

Рис. 1.13. Изменение возбудимости в различные фазы потенциала действия:
1 — исходное состояние; 2— начальное повышение возбудимости; 3 — абсолютная рефрактерная фаза; 4 — относительная рефрактерная фаза; 5 — фаза экзальтации; 6 — вторичное снижение возбудимости
Изменение возбудимости в динамике одиночного цикла возбуждения. В динамике развертывания одиночного цикла возбуждения происходят изменения возбудимости ткани. Это определяется путем нанесения на ткань вслед за первым (кондиционирующим) стимулом через различные интервалы времени второго (тестирующего) стимула. Установлено (рис. 1.13), что местному потенциалу соответствует некоторое повышение возбудимости. Спайковому потенциалу соответствует абсолютная рефрактерная фаза. С началом быстрой реполяризации наблюдается относительная рефрактер- ность. Отрицательный следовой потенциал характеризуется повышением возбудимости.
Завершается одиночный цикл возбуждения повторным снижением возбудимости — периодом субнормальной возбудимости, совпадающим с положительным следовым потенциалом и развитием гиперполяризации мембраны.
Продолжительность абсолютной рефрактерной фазы определяет функциональную лабильность соответствующей ткани и препятствует суммации последующих возбуждений. В среднем для исчерченного скелетного мышечного волокна период абсолютной рефрактерности равен 2,5—4,0 мс, для толстых нервных волокон — 0,4 мс.
1.4. ТОРМОЖЕНИЕ
Торможение — активный биологический процесс, который препятствует и предотвращает развитие возбуждения либо сменяет, прекращает уже развившийся процесс возбуждения.
Процесс торможения — активный процесс, связанный с большими энергозатратами.
Тормозные влияния нервной системы на деятельность сердца были впервые открыты в 1847 г. братьями Вебер при раздражении блуждающих нервов (см. разд. 5.1.6).
Существует две точки зрения на происхождение торможения:
• торможение как самостоятельный биологический процесс;
• торможение как результат взаимодействия двух возбуждений. Последняя точка зрения берет свое начало от И. М. Сеченова.
Электрофизиологические проявления торможения. Современные электрофизиологические исследования показывают, что в
основе процесса торможения лежит гиперполяризация мембраны возбудимой ткани. При этом повышается проницаемость калиевых каналов и каналов для ионов СГ. В результате регистрируемый на мембране потенциал приобретает обратную (по направленности) спайковому процессу форму (рис. 1.14). В синаптической области тормозные влияния соответствующих медиаторов проявляются на постсинаптической мембране в форме тормозных постсинаптических потенциалов (ТПСП).

Рис. 1.14. Тормозной постсинаптический ответ нейрона на песси- мальное раздражение (обозначено стрелкой)
Классификация торможения. По локализации различают торможение:
• периферическое;
• центральное.
Примером периферического торможения является тормозное действие блуждающего нерва на сердце, адреналина — на моторную деятельность кишечника и др.
Центральное торможение открыто И. М. Сеченовым (1862). Оно получило название сеченовского торможения. Центральное торможение может быть спинальным, ретикулярным, корковым (см. разд. 3.4.2).
Природа торможения. Вопрос о природе торможения до сих пор окончательно не решен.
Н. Е. Введенский связывал процесс торможения с состоянием пессимума и даже парабиоза тканей (пессимальное торможение). При парабиозе, по существу, происходит процесс взаимодействия двух возбуждений. Одно из них исходное, статическое, «застойное», а другое — кондиционирующее, тестирующее, которое не проходит через парабиотический участок.
Как указывалось выше, гиперполяризация развивается после возбуждения как следовое торможение.
В основе процесса торможения лежат:
• устойчивая деполяризация;
• гиперполяризация;
• инертная поляризация (трудно возбуждаемая ткань) и абсолютная рефрактерность.
Возможно, природа всех этих состояний едина.
Тормозные химические факторы. В нервных окончаниях выделяются специальные биологически активные вещества (медиаторы и модуляторы), некоторые из них — например, у-аминомасляная кислота (ГАМК), глицин и др. — способны вызывать гиперполяризацию мембран нервных клеток и торможение их активности.
1.5. СОКРАЩЕНИЕ
Сокращение возникло в эволюции живых существ как необходимая функция, прежде всего — перемещения внутренних органелл и самих субъектов в пространстве, включая движения органов чувств. Сократительные функции направлены на удовлетворение ведущих потребностей живых существ. Среди разнообразных форм сокращений ведущая роль принадлежит мышечным сокращениям.
Мышцы подразделяются на скелетные и гладкие.
Скелетные мышцы. Главной особенностью скелетных мышц является то, что они в живых организмах подчиняются произвольному контролю.
Миофибриллы, составляющие скелетные мышцы, подразделяются на саркомеры, включающие темные в обычном свете анизотропные диски (A-диски) с сильным двойным лучепреломлением и изотропные диски (1-диски), имеющие слабое двойное лучепреломление, что создает картину поперечной исчерченности мышечного волокна (рис. 1.15).
Сокращения скелетных мышц подразделяются на фазные и тонические.
Быстрая (фазная) активность скелетных мышц определяется белыми мышечными волокнами, которые отличаются значительным содержанием миофибрилл, гликогена и гликолитических ферментов. Гликоген обеспечивает энергетические потребности мышцы.

|
Тонические сокращения определяются красными мышечными волокнами. В них содержится большое количество миоглобина и цитоплазматических органелл, особенно митохондрий.
Красные мышечные волокна способны к длительному напряжению и к более длительной динамической работе. Белые волокна, миофибриллы которых толще и сильнее, быстро сокращаются, но и легко утомляются. Большинство мышц — смешанные, они в разных пропорциях содержат белые и красные волокна.
Разновидность мышечного сокращения. Различают изотонический, изометрический и ауксо- тонический режимы мышечного сокращения. При изотоническом сокращении напряжение мышцы при ее возбуждении не изменяется, изменяется лишь ее длина, как в случае передвижения груза. При изометрическом сокращении изменяется напряжение мышцы при неизменной ее длине (например, при удержании груза в одном положении). При ауксотоническом режиме изменяются напряжение и длина мышц, что наблюдается при различных видах деятельности.
Общебиологические свойства мышц:
• Обеспечение перемещения тела и его отдельных частей в пространстве для удовлетворения ведущих потребностей организма.
• Антигравитационные тонические функции, поддержание позы.
• Участие в терморегуляции (мышечная дрожь).
• Участие в регуляции внутренней среды (перемещение пищевых веществ, крови, лимфы и мочи, депонирование углеводов и жировых веществ и др.).
• Антистрессорные влияния.
• Тоническая «подпитка» функций мозга.
• Биоэлектрическая активность (общий смысл ее пока еще не совсем ясен).
Физиологические свойства скелетных мышц:
• Возбудимость.
• Проводимость.
• Сократимость.
• Эластичность.
Возбудимость мышц. В ответ на действие адекватных раздражителей в мышцах формируется распространяющийся процесс возбуждения. Возбуждение мышц осуществляется прямым способом, когда раздражитель действует непосредственно на мышцу, и непрямым способом — через первичное раздражение иннервирующего мышцу нерва. Мерой возбудимости мышцы является порог раздражения, хронаксия, аккомодация, функциональная лабильность (см. разд. 1.2).
При действии на мышечное волокно слабых, подпороговых раздражений в нем возникает местный процесс возбуждения. Только при увеличении силы раздражения и достижении критической точки возникает распространяющийся по мышце процесс возбуждения. Таким образом, скелетные мышцы, как и другие возбудимые ткани, возбуждаются по триггерному механизму.
Важным физиологическим свойством скелетной мышцы является изменение ее возбудимости в процессе возбуждения (см. ниже).
Проводимость — проведение возбуждения по мышечному волокну.
Сократимость — способность мышцы сокращаться в ответ на возбуждение.
При градуальном увеличении силы раздражения целая мышца постепенно увеличивает до определенной величины амплитуду сокращений за счет того, что различные составляющие ее волокна имеют различную возбудимость.
Одиночное мышечное волокно подчиняется закону «все или ничего», т.е. уже на пороговое воздействие мышца дает максимальное по амплитуде сокращение.
Если учесть, что на подпороговые раздражители в мышце возникает местный процесс возбуждения, то этот закон приобретает форму «все или кое-что».
При достаточной силе и частоте раздражения мышца снижает амплитуду сокращения. В ней развивается состояние пессимума.
Эластичность — способность мышцы после растяжения восстанавливать исходную длину.
Одиночное мышечное сокращение. В ответ на одиночное раздражение скелетная мышца отвечает одиночным сокращением (рис. 1.16).
 Рис. 1.16. Одиночное мышечное сокращение (механическая запись):
Рис. 1.16. Одиночное мышечное сокращение (механическая запись):
А — сокращеннию предшествует потенциал действия (1 — латентный период; 2 — фаза сокращения; 3 — фаза расслабления); Б — изменения возбудимости мышцы
Одиночное мышечное сокращение включает латентный период, фазу сокращения и расслабления.
Изменения возбудимости скелетной мышцы в процессе одиночного цикла сокращения. В процессе одиночного цикла сокращения динамически изменяется возбудимость мышцы. В латентный период, когда в мышце возникает потенциал действия, наблюдаются описанные выше фазы изменения возбудимости: абсолютная рефрактерная фаза, относительная рефрактерность и экзальтационная фаза (см. рис. 1.16, Б).
Суммация мышечных сокращений. Мышечное волокно обладает способностью к суммации (суперпозиции) сокращений. Если на мышцу подаются два раздражающих импульса, причем второй импульс попадает в фазу повышенной возбудимости от предыдущего раздражения, амплитуда второго мышечного сокращения значительно возрастает за счет суммации.
Реакция мышцы на ритмическое раздражение. При ритмическом раздражении, если повторные раздражения осуществляются в фазу расслабления от предыдущих раздражений, происходит сложение (суммация) сокращений и возникает зубчатый тетанус. Если повторное раздражение попадает в фазу сокращения от предыдущего раздражения, возникает гладкий тетанус. При еще большей частоте раздражений, если последующее раздражение попадает в фазу латентного периода, особенно рефрактерную фазу возбудимости предшествующего раздражения, мышца перестает отвечать на повторные раздражения. Возникает пессимум раздражения (рис. 1.17).

ОНШМНШШ------ МЯНМНИ1
Рис. 1.17. Суммация одиночных сокращений поперечно-полосатой мышцы в зависимости от начала 2-го раздражения и образование тетануса:
1 — ответ на одиночные раздражения; 2 — зубчатый тетанус; 3 — гладкий тетанус; 4 — пессимум сокращения мышцы при еще большем увеличении частоты раздражения
Заметим, что амплитуда гладкого тетануса больше зубчатого и одиночного сокращения, так как при этом последующее раздражение попадает в фазу экзальтации от предшествующих раздражений.
Теории тетануса:
• Механическая теория суперпозиции сокращений (Г. Гельмгольц). При этом каждое последующее сокращение формируется как бы с исходного уровня.
• Теория изменения состояния мышцы при воздействиях (Н. Е. Введенский), согласно которой предыдущее состояние мышцы определяет ее ответ на последующее раздражение.
Следует иметь в виду, что тетанус определяет только плавность движений, удержание предмета в руке. Но это дорогой в энергетическом плане процесс. При этом быстро развивается утомление.
Нейромоторные единицы. Установлено, что каждое отдельное нервное волокно иннервирует несколько (от 10 до 3 тыс.) мышечных волокон. При этом каждое отдельное мышечное волокно может иннервироваться от нескольких различных нервных волокон. Чем меньшее количество мышечных волокон иннервируется одним нервным волокном, тем точнее движение.
В различных мышцах количество мышечных волокон в нейро- моторной единице различно. В наружных мышцах глазного яблока, например, их 3—6, в пальцах руки — 10—25, а в икроножной мышце нижних конечностей — до 7 тыс.
Внутри нейромоторных единиц за счет блокады отдельных нейромышечных синапсов происходит асинхронное чередующееся сокращение мышечных волокон. Только при крайних мышечных усилиях мышечные волокна начинают функционировать синхронно вместе с соответствующими нервными центрами.
Тоническая активность скелетных мышц. Специальные структуры мозга, в частности экстрапирамидная система (см. разд. 3.14 и 7.2), обеспечивают тоническую активность скелетных мышц, позу тела и грубые движения. Моторные единицы экстрапирамидной системы охватывают до 2—3 тыс. мышечных волокон.
Мышечная деятельность — результат взаимодействия тетанических, асинхронных и тонических сокращений мышечных волокон. При мышечной тренировке создаются определенные соотношения различных комбинаций разнообразных сокращений мышечных волокон, обеспечивающих достижение необходимых для организма полезных приспособительных результатов. Отсюда понятна беспредельность спортивных рекордов.
Оптимальная частота в работе мышц. И. М. Сеченов считал, что оптимальная частота мышечной деятельности, определяющая оптимальную работоспособность человека, составляет:
Сокращение _ 1
Расслабление 3
Оптимальное сокращение мышц может быть достигнуто при применении электростимуляторов с обратной связью от показателя биоэлектрической активности мышц. В этом случае, как только мышца утомляется, стимулятор прекращает воздействие. Такие стимуляторы широко применяются в спортивной практике и при лечении двигательных параличей.
Мышечное утомление. При длительном действии на мышцу частых раздражений, даже в пределах оптимума раздражений, наблюдается постепенное удлинение всех фаз мышечных сокращений. Мышца сначала переходит в состояние зубчатого тетануса, а затем — в состояние утомления. При этом, несмотря на отсутствие сокращений, в мышце все еще регистрируются биопотенциалы (рис. 1.18).

Рис. 1.18. Динамика утомления поперечно-полосатой мышцы при ее длительном ритмическом раздражении (7-5— изменение формы одиночного мышечного сокращения по мере утомления мышцы)
Теории мышечного утомления: теория накопления продуктов обмена, в частности молочной и фосфатной кислот (Э. Пфлюгер); теория истощения депо кальция; теория истощения энергетических запасов, в частности гликогена и АТФ (М. Шифф).
В целом организме утомляются сначала нервные центры, затем синапсы и только после этого мышечные волокна.
Гладкие мышцы. Образуют стенки желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других внутренних органов. Мультиунитарные гладкие мышцы в ресничной мышце глаза, мигательной перепонке, в кровеносных сосудах, в волосяных клетках и т.п. функционируют изолированно, независимо друг от друга.
Эти мышцы иннервируются, как правило, отдельными нервными окончаниями.
У унитарных гладких мышц мембраны отдельных волокон сливаются посредством электрических контактов (нексусов), которые имеют низкое электрическое сопротивление. За счет этого электрические потенциалы распространяются с одного мышечного волокна на другое. В результате унитарные гладкие мышцы представляют собой функциональный синцитий. Такие мышцы составляют пищеварительный тракт, матку и мочеточники.
По сравнению со скелетными мышцами гладкие мышцы обладают рядом особенностей:
• обладают более низкой возбудимостью;
• одиночный цикл сокращения у них растянут до 1 мин за счет удлинения всех фаз: латентного периода, сокращения и расслабления;
• обладают меньшей функциональной лабильностью и большей хронаксией; способны давать тетанус при редкой частоте раздражений; при увеличении частоты раздражений легко впадают в состояние пессимума;
• обладают медленным развитием утомления;
• характеризуются длительными тоническими сокращениями и относительно медленными спонтанными ритмическими движениями;
• обладают пластическим тонусом: под влиянием нагрузки изменяют длину, теряют напряжение и длительно без больших затрат энергии сохраняют это состояние; после прекращения нагрузки не сразу возвращаются к исходной длине (пример: растянутый пищей желудок или растянутый мочой мочевой пузырь);
• обладают запирательными свойствами (особенно мышцы сфинктеров), длительно удерживаются в сокращенном состоянии без больших энергетических затрат;
• в отличие от скелетных мышц, которые в ответ на действие постоянного тока дают единичной ответ и затем переходят в состояние аккомодации, гладкие мышцы при действии постоянного тока проявляют ритмические сокращения (так называемые итеративные системы)',
• некоторые гладкие мышцы (например, кишечника) способны к автоматии, т.е. спонтанным сокращениям без видимой внешней причины;
• высокочувствительны к химическим веществам, в частности медиаторам, олигопептидам и гормонам (например, ацетилхолин вызывает сокращение гладких мышц кишечника в концентрации 1 х 10 s);
• обладают способностью к псевдотетаническим сокращениям: в ответ на одиночное раздражение проявляют тетанический ответ, так как отвечают на каждую порцию выделяющегося при этом ацетилхолина.
Механизм мышечного сокращения. Процесс сокращения мышцы связан с изменением длины саркомеров. Изменение длины саркомеров обусловлено взаимным перекрытием актиновых и миозиновых филаментов.
Строение саркомеров. Каждый саркомер ограничен с боков Z-днсками. Основными компонентами саркомеров являются белковые молекулы актина и миозина. Каждая нить актина образована двумя перекрещивающимися цепями мономеров. В углублениях между цепочками мономеров актина располагаются нити белка тропомиозина. На регулярных интервалах этих нитей (примерно через 40 нм) располагаются молекулы белка тропонина. Тропониновые молекулы состоят из тропомиозинсвязывающих (ТпТ), кальцийсвязывающих (ТпС) и тормозных (Tnl) субъединиц.
Нити актина состоят из множества собранных в пучки молекул, каждая из которых имеет стержневой компонент и отходящие от них шейку и головки (рис. 1.19).
В настоящее время наиболее распространенной теорией, объясняющей процесс сокращения мышцы, является теория скольжения нитей.
Теория скольжения нитей. При поступлении нервных импульсов к мышце происходит деполяризация мембраны концевой пластинки нервного окончания. Потенциал действия по плазматической мембране миофибриллы быстро распространяется по продольным и поперечным Т-трубочкам. Благодаря этому сигнал передается эндоплазматической сети. В результате из концевых и латеральных цистерн саркоплазматической сети освобождаются ионы Са;\ Ионы Са2+ связываются с субъединицами тропонина — тропонином С.
Процесс, при котором деполяризация мышечных волокон вызывает их сокращение, получил название сопряжение возбуждения и сокращения.
В исходном состоянии взаимодействие между актином и миозином тормозится субъединицей тропонина I. В результате связы-
 |
|
 |
| |||||
| |||||
| |||||
 | |||
 | |||
|
 |
|
вания ионов Са2; с субъединицей тропонина С тормозящее влияние тропонина I на актин ослабевает, что позволяет тропомиозину смещаться латерально (рис. 1.20). Это освобождает связывающие места для прикрепления миозиновой головки. Головка миозиновой молекулы, прикрепляясь к связывающей зоне молекулы актина, поворачивается под углом в 90°, вызывая тем самым скольжение
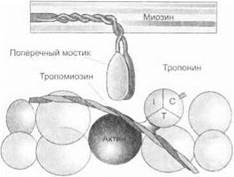
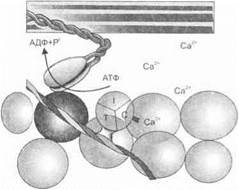
Рис. 1.20. Механизм мышечного сокращения
нити актина. Затем происходит отсоединение головки миозина и последующее ее прикрепление к следующему месту связывания. В головках миозина содержится фермент аденозинтрифосфатаза (АТФаза), который при контакте головки миозина с актиновой молекулой осуществляет гидролиз фосфатных связей АТФ. Каждая толстая нить миозина имеет около 500 головок, и каждая за время быстрого сокращения мышцы осуществляет около пяти раз циклы взаимодействия с актином.
Вскоре после освобождения ионов Са2+ саркоплазматическая сеть начинает реаккумулировать их в продольные канальцы. Отсюда они диффундируют в терминальные цистерны, где хранятся до следующих процессов возбуждения. В то же время ионы Са2+,
находящиеся вне саркоплазматической сети, существенно снижают взаимодействие между миозином и актином и способствуют тем самым расслаблению мышцы. АТФ обеспечивает энергию для активного транспорта кальция в саркоплазматическую сеть.
Кровоснабжение мышцы. Все мышцы пронизаны сетью кровеносных сосудов, особенно красные волокна. Капилляры скелетных мышц — короткие. Скорость передвижения крови в них незначительна. При сокращении скелетных мышц приток крови к ним замедляется, венозный отток ускоряется. В этом проявляется работа так называемого мышечного насоса. При мышечной работе дополнительно раскрываются длинные петли кровеносных капилляров.
Энергетические процессы мышечного сокращения. Процесс мышечного сокращения — активный процесс. Он тесно связан с метаболическими энергетическими процессами.
Единственным источником энергии для сокращения мышц служит АТФ. Запасы АТФ в мышце незначительны. В мышце имеется три источника образования АТФ: распад креатинфосфата (КФ), анаэробный гликолиз и окислительное фосфорилирование субстратов в митохондриях. АТФ и КФ — соединения нестойкие, они расщепляются с образованием богатой энергетическими связями молекулы фосфора:
АТФ -> АДФ + Р', КФ -> К + Р'.
Выявлено, что при работе мышцы содержание в ней АТФ и КФ не изменяется. Наряду с распадом этих веществ идет их синтез. Источником ресинтеза АТФ и КФ в мышцах является гликоген.
Установлено, что работающая мышца потребляет значительное количество кислорода. В атмосфере чистого азота изолированная мышца может работать, но непродолжительно.
Химический ресинтез АТФ осуществляется в работающих мышцах анаэробным (без участия кислорода) и аэробным (с участием кислорода) путем. Начальная анаэробная фаза мышечного сокращения осуществляется за счет энергии содержащихся в мышцах креатинфосфата и гликогена.
Под влиянием гликолитических ферментов в анаэробных условиях (например, при длительной работе мышц) энергия освобождается в процессе расщепления углеводов (гликогена и глюкозы) до молочной кислоты.
Анаэробные процессы не обеспечивают длительно мышечное сокращение. Необходим кислород, под влиянием которого молочная кислота расщепляется до молекул воды и углекислого газа. При этом выделяется дополнительная энергия, определяющая распад молекул гликогена. Без достаточного поступления кислорода в мышцах в результате накопления молочной кислоты наблюдается так называемая кислородная задолженность (кислородный долг).
Особенности энергетического обеспечения быстрых и медленных мышечных волокон. Быстрые и медленные мышечные волокна имеют разный тип энергетического обеспечения. Быстрые мышечные волокна, богатые гликогеном, работают преимущественно на энергетической основе анаэробной фазы мышечного сокращения. Они быстро утомляются. Волокна медленных мышц, богатые митохондриями и миоглобином, при работе используют в основном энергию аэробных реакций. Они хорошо кровоснабжаются и могут долго работать без утомления.
При работе большой интенсивности в мышцах окисляются в основном углеводы, а при малой интенсивной работе — свободные жирные кислоты.
Теплообразование в мышце. Мышца не является тепловой машиной. В ней химическая энергия превращается в механическую. При этом выделяется тепло. В мышце наблюдается обусловленное метаболическими процессами теплообразование покоя. Особенности химических реакций находят отражение в теплообразовании мышцей (А. Хилл). В фазу сокращения наблюдается начальное теплообразование и теплообразование, связанное с укорочением мышцы. После сокращения при расслаблении в мышце также образуется тепло, по количеству равное наблюдаемому при сокращении (конечное, отставленное теплообразование).
1.6. СЕКРЕЦИЯ
Элементарным процессом жизнедеятельности является процесс секреции.
Секреция — процесс образования в клетках и последующего выделения специфического продукта — секрета.
Различают внутреннюю и внешнюю секрецию. При внутренней секреции секрет выделяется во внутреннюю среду организма —
кровь, тканевую жидкость и лимфу. Клетки, участвующие во внутренней секреции, называются эндокринными.
При внешней секреции секрет выделяется на поверхность слизистой оболочки, кожи или в просвет пищеварительного тракта. Этот вид секреции получил название экзокринной.
Секреторные клетки выделяют самые различные по структуре и свойствам вещества: белки, гликозаминогликаны, липиды, ферменты, растворы оснований, солей, кислот и воду.
Секретируемые вещества выступают в качестве: а) собственных секретов; б) экскретов; в) рекретов.
Секреты — продукты клеточного анаболизма. Их синтез и выделение являются основной функцией секреторной клетки.
Экскреты — конечные продукты клеточного катаболизма; как правило, подлежат выделению из организма.
Рекреты — вещества, которые сначала поглощаются клеткой, а затем выделяются ею в неизменном виде.
Биоэлектрические свойства секреторных клеток. Возбуждение секреторных клеток сопровождается дискретными изменениями поляризованное™ их мембран — секреторными потенциалами. Возбуждение одних секреторных клеток связано с их деполяризацией (экзокринные клетки поджелудочной железы, клетки протоков слюнных желез). Возбуждение других секреторных клеток приводит к процессам гиперполяризации.
Базальная и апикальная мембраны секреторных клеток характеризуются разной степенью поляризации. За счет разности потенциалов этих мембран создается электрическое поле напряжением 20—30 мВ, которое удваивается при возбуждении клетки. Электрическое поле способствует перемещению зрелых гранул секрета к апикальному полюсу клетки, взаимодействию их с мембраной и выходу их из клетки путем диффузии и экзоцитоза.
Секреторный цикл излагается в курсе гистологии.
Регуляция секреции отдельных желез будет рассмотрена в соответствующих разделах.
1.7. РЕЦЕПЦИЯ
Рецепция — процесс восприятия и трансформации (преобразования) механической, термической, электромагнитной и химической
энергии в сложную последовательность мембранных и цитоплазматических процессов, порождающих нервные сигналы.
Рецепторы подразделяются на клеточные (молекулярные) и окончания нервов (сенсорные).
Клеточная рецепция. Роль клеточных рецепторов выполняют пронизывающие мембраны клеток или находящиеся на их поверхности белковые молекулы.
Белковые рецепторы мембран по принципу «ключ-замок» связывают специальные, соответствующие их молекулярной структуре вещества, называемые лигандами. Взаимодействие рецепторов с лигандами через активные центры связывания составляют процессы клеточного узнавания.
Рецепторы локализуются не только на поверхности, но и внутри клетки — в цитоплазме и ядре.
Белки рецепторов постоянно синтезируются в ядрышках ядер и после сложных взаимодействий с транспортной и информационной РНК поступают в цитоплазму. Дальнейшее созревание, формирование и транспорт клеточных рецепторов происходит в гранулярной эндоплазматической сети с поглощением энергии окружающих комплекс Гольджи многочисленных митохондрий.
После связывания с лигандом рецептор передает информационный сигнал на исполнительные образования клетки: открывает или закрывает ионный канал, активирует специфические ферменты или действует сам в качестве ионофора.
Рецепторы для веществ белковой и пептидной природы расположены на наружной поверхности клеточных мембран. Рецепторы стероидных гормонов и производных тирозина расположены в цитоплазме и ядре клеток.
Мембранная рецепция. При взаимодействии лиганда с мембранным рецептором активируется каскад внутриклеточных ферментативных реакций. Возбуждается мембранная аденилатциклаза. Взаимодействуя с АТФ, мембранная аденилатциклаза стимулирует образование циклического аденозинмонофосфата (цАМФ). цАМФ стимулирует внутриклеточные ферментные системы — протеин- киназы, активирующие геном клеток, и способствует тем самым синтезу специфических клеточных белков.
Ядерная рецепция. Стероидные и тиреоидные гормоны, растворимые в лигандном слое клеточных мембран, свободно проходят
в цитоплазму, где они соединяются со специфическими рецепторными белками. После ряда превращений комплекс «лиганд-рецептор» проникает в ядро и реализует свое действие на генетическом уровне.
Сенсорная рецепция представлена окончаниями нервных клеток. Болевые рецепторы представлены свободными нервными окончаниями. Обонятельные — чувствительными нейронами. Вкусовые — эпителиальными клетками, связанными с помощью синапсов с афферентными нейронами. Рецепторы кожи бывают разных видов (рис. 1.21). Сложноорганизованными рецепторными органами являются органы зрения, слуха и равновесия (см. гл. 6).
Сенсорная рецепция осуществляется с помощью специальных, находящихся в окончаниях нервов рецепторных белков, взаимодействие которых с соответствующими раздражителями приводит к изменению ионной проводимости мембраны и возникновению локального рецепторного потенциала.
Стимуляция сенсорных рецепторов в живых организмах осуществляется такими естественными раздражителями, как свет, звук, давление, температура, кислотность среды, электромагнитные волны и т.п.
Рецепторный потенциал характеризуется тем, что под влиянием раздражителя в рецепторе сначала до критического уровня возрастает возбудимость, отражением чего служит местный процесс возбуждения. При достижении критического уровня изменения возбудимости начинается процесс генерации распространяющихся потенциалов.
Различают первично- и вторичночувствующие рецепторы.
Первичночувствующие рецепторы. В первичночувствующих рецепторах стимул действует непосредственно на воспринимающий субстрат (рецепторный белок), заложенный в самой воспринимающей биполярной рецепторной клетке. При этом в клетке в результате изменения проницаемости мембраны к ионам Na1 или Са2+ возникает рецепторный (местный) потенциал. Рецепторный потенциал электротонически распространяется по аксонам воспринимающих биполярных (сенсорных) клеток.
К первичночувствующим рецепторам относятся: обонятельные, тактильные, температурные рецепторы и мышечные веретена.
 |
| ||||
| ||||
 |
| ||||||
| ||||||
| ||||||
 |
|
 |
|
Рис. 1.21. Разновидности рецепторов кожи
На рис. 1.22 представлен первичночувствующий рецептор растяжения рака. Он состоит из крупной нервной клетки с аксоном и дендритами. Рецепторные дендриты нервной клетки контактируют с мышечным волокном. При растяжении мышечных волокон и расположенных на них дендритах происходит деполяризация мем
браны рецепторной клетки. В клетке регистрируется рецепторный потенциал. Деполяризация исчезает с прекращением стимуляции. Длительность рецепторного потенциала соответствует длительности, а его амплитуда возрастает с увеличением интенсивности раздражающего стимула. Рецепторный потенциал в рецепторе растяжения обусловлен повышением натриевой проводимости мембраны растянутых дендритов. Рецепторный потенциал электротонически распространяется по всей рецепторной клетке.

Рис. 1.22. Рецепторный потенциал и потенциалы действия в рецепторе растяжения:
А — схематическое изображение рецептора растяжения рака; стимул деполяризует дендриты, вызывая рецепторный потенциал, который регистрируется в соме; Б — ритмические потенциалы действия, которые возникают в основании аксона под влиянием рецепторного потенциала (а, б, в, г — местные потенциалы; д — распространяющийся потенциал)
В отличие от рецепторной клетки, в ответ на раздражение в аксоне первичночувствующих рецепторов возникает серия потенциалов действия. Под влиянием рецепторного потенциала в аксоне
рецепторной клетки формируется генераторный потенциал — импульсный процесс распространения возбуждения (рис. 1.23). Частота импульсаций аксонов зависит от амплитуды рецепторного потенциала. Трансформация рецепторного потенциала в серию потенциалов действия во многих рецепторах происходит около места, где аксон отходит от рецепторной клетки.
Рис. 1.23. Генераторный потенциал вторичночувствующе- го рецептора (по А. Гайтону, 1985):
РП — рецепторный потенциал; МП — мембранный потенциал
 Вторичночувствующие рецепторы. У этих рецепторов между действующим стимулом и воспринимающим сенсорным нейроном располагаются дополнительные рецептирующие клетки, как правило, не нервного происхождения (например: колбочки и палочки в сетчатке). Между рецептивной и сенсорной клеткой имеются синапсы. При возбуждении рецептирующих клеток в них формируются локальные рецепторные потенциалы, под влиянием которых в синапсах выделяется медиатор, возбуждающий сенсорные нейроны. В сенсорных нейронах возникает генераторный потенциал, который сначала электротонически распространяется по нервным волокнам. В электрогенных участках нервных волокон генерируются потенциалы действия.
Вторичночувствующие рецепторы. У этих рецепторов между действующим стимулом и воспринимающим сенсорным нейроном располагаются дополнительные рецептирующие клетки, как правило, не нервного происхождения (например: колбочки и палочки в сетчатке). Между рецептивной и сенсорной клеткой имеются синапсы. При возбуждении рецептирующих клеток в них формируются локальные рецепторные потенциалы, под влиянием которых в синапсах выделяется медиатор, возбуждающий сенсорные нейроны. В сенсорных нейронах возникает генераторный потенциал, который сначала электротонически распространяется по нервным волокнам. В электрогенных участках нервных волокон генерируются потенциалы действия.
К вторичночувствующим относятся рецепторы слуха, зрения, вкуса, вестибулярные.
Вторичночувствующие слуховые, вестибулярные клетки представлены волосковыми клетками. Апикальная поверхность волосковой клетки, находящаяся в межклеточной жидкости, имеет
пучок волосков. Смещение волосков в одну сторону деполяризует рецептивную клетку, а в другую — ее гиперполяризует. При этом изменяется ионная проводимость мембраны клетки. В покое клетка имеет небольшое количество открытых калиевых каналов. Смешение волосков мембраны приводит к усилению входящего потока ионов Na'.
Классификация рецепторов. В соответствии с модальностью ощущения выделяют зрительные, слуховые, осязательные, обонятельные, вкусовые, холодовые, тепловые и болевые рецепторы.
По локализации выделяют экстерорецепторы и интерорецеп- торы. К экстерорецепторам относятся фоторецепторы, слуховые, тактильные, температурные и хеморецепторы, расположенные на поверхности тела и в начальных отделах пищеварительного тракта и дыхательных путей.
К интерорецепторам относятся рецепторы внутренних органов, сосудов, опорно-двигательного аппарата — мышц, сухожилий, костей и суставов.
Рецепторы подразделяют на дистантные и контактные. Первые воспринимают информацию от источника, расположенного на некотором расстоянии от них (зрительные, слуховые), а вторые — при непосредственном соприкосновении с раздражителями (тактильные).
Однако такое деление не вполне правомерно, поскольку на самом деле ощущения и света, и звука возникают вследствие непосредственного воздействия электромагнитных колебаний на соответствующие рецепторные аппараты сетчатки и кортиева органа.
Структура рецепторов. Рецепторы могут быть представлены свободными нервными окончаниями; окончаниями, покрытыми особой капсулой (инкапсулированные); иметь вид палочек, колбочек, ветвей, гроздьев, щеточек, волосков и др. Некоторые рецепторы объединяются в сложноорганизованные множества — сетчатку глаза, кортиев орган внутреннего уха и др.
Специализация рецепторов. Характерным свойством рецепторов является их высокая генетически детерминированная специализация к восприятию адекватных раздражителей. В соответствии с природой адекватных раздражителей выделяются:
• тактильные рецепторы кожи;
• фоторецепторы сетчатки — нервные элементы, возбуждаемые электромагнитными волнами длиной 300—900 нм;
• слуховые, вестибулярные и гравитационные рецепторы внутреннего уха;
• рецепторы опорно-двигательного аппарата (растяжения, суставные, сухожильные, мышц);
• барорецепторы сердца и сосудов;
• хеморецепторы каротидных телец, обоняния, вкуса, а также кровеносных сосудов и тканей, реагирующих на изменение химического состава жидких внутренних сред организма;
• терморецепторы кожи, внутренних органов и центральной нервной системы (ЦНС), реагирующие на изменения температуры окружающей среды и внутренней среды организма.
Кроме этого выделяют рецепторы вибрации, рецепторы волосяных фолликулов, ганглиев и многие другие.
Модальность. Некоторые (мономодальные) рецепторы приспособлены для восприятия лишь одного вида раздражения — например, вкусовые рецепторы сладкого; другие (полимодальные) — для восприятия нескольких видов раздражителей, например, ноцицеп- торы кожи, участвующие в формировании болевого ощущения при любом механическом, химическом и температурном повреждающем воздействии.
Чувствительность. Большинство рецепторов обладает высокой чувствительностью по отношению к адекватным раздражителям. Вместе с тем выделяют низко- и высокопороговые рецепторы.
Низкопороговые — наиболее чувствительные рецепторы расположены в коже (тактильные, или осязательные, волоски), в сетчатке глаза (палочки), в обонятельных луковицах. Так, достаточно всего 1 кванта света, чтобы вызвать возбуждение фоторецептора, отвечающего за ахроматическое (черно-белое) зрение, и 7 квантов — чтобы возникло светоощущение. Хорошо известна исключительно высокая чувствительность некоторых животных к половым феромонам, выделяемым особями противоположного пола и распознаваемым на огромных расстояниях. У этих особей иногда достаточно всего нескольких молекул пахучего вещества, чтобы возбудить обонятельные рецепторы и вызвать ощущение запаха.
К высокопороговым — наименее чувствительным — относятся рецепторы сетчатки (колбочки), ответственные за хроматическое (цветовое) зрение, и ноцицепторы кожи, возбуждающиеся при механическом воздействии большой интенсивности.
Самые малые значения раздражителей, способные вызвать ощущение, называются порогами чувствительности, или абсолютными порогами.
Закон специфической энергии. Некоторые рецепторы возбуждаются и неадекватными раздражителями. Например, умеренное механическое раздражение сетчатки (надавливание на глазное яблоко через закрытое веко) вызывает ощущение света (фосфен). Сильное механическое раздражение (например, удар по глазу) сопровождается субъективно переживаемым ощущением «искр».
И. Мюллером сформулирован закон специфической энергии. Закон гласит: восприятие зависит не от раздражителя, а от той внутренней энергии, которая выделяется в рецепторах при их раздражении. Закон отражает субъективную сторону восприятия разнообразных воздействий.
Особенности передачи информации от рецепторов в ЦНС. Все больше данных свидетельствует о том, что в живых организмах информация от рецепторов в ЦНС передается в форме частотно- модулируемых электромагнитных волн.
Данный способ «частотно-амплитудной модуляции» обнаружен у многих рецепторов: кожных рецепторов давления, у мышечных веретен, в барорецепторах артерий, в фоторецепторах сетчатки и др. — все они преобразуют интенсивность раздражающих стимулов в среднюю частоту импульсаций.
Частота нервных импульсов является универсальным носителем информации.
Первичная трансформация стимула в рецепторный потенциал обычно называется преобразованием. Рецепторы можно рассматривать в качестве первичных преобразователей информации.
Характерно, что стимул не является источником энергии для рецепторного потенциала. Он только контролирует движение ионов (в частности, ионов Na+) через мембрану, основанное на трансмембранной разности их концентраций. Один квант света уже может вызвать такие значительные мембранные токи, что появляющийся рецепторный потенциал инициирует заметные изменения активности зрительной клетки.
Такой тип ответных реакций, как указывалось ранее, получил в живых тканях название триггерных. В триггерных механизмах ответ возбудимой ткани определяется не столько энергией воздействую-
шего раздражителя, сколько сложными молекулярными изменениями, происходящими при действии соответствующих стимулов на молекулярные рецепторы мембран. При этом ответ рецептора формируется только в том случае, когда его возбудимость, т.е. молекулярные трансформации, достигает определенного критического уровня. Триггерные механизмы представляют одну из форм информационных преобразований в живых организмах, строящихся на молекулярных соотношениях воздействия стимулов и активной реакции живой ткани. При этом реакция ткани осуществляется без потери информационного смысла раздражения.
Адаптация рецепторов. В большинстве рецепторов рецепторный потенциал во время постоянной стимуляции падает или адаптируется. Разные рецепторы сильно отличаются по скорости адаптации. Рецепторный потенциал рецептора растяжения адаптируется относительно слабо. Медленно адаптируются рецепторы, контролирующие степень растяжения мышц, хеморецепторы — к содержанию ионов Н+ в тканях, осморецепторы. Другие рецепторы, такие как, например, терморецепторы кожи и световые рецепторы, адаптируются с промежуточной скоростью. Быстро адаптируются рецепторы давления и вибрации (тельца Пачини).
Адаптация происходит не только на уровне рецепторов, но и на нейронах ЦНС, к которым от рецепторов поступает информация. Адаптация тактильных рецепторов проявляется также в снижении частоты импульсации в чувствительных волокнах, несмотря на не- прекращающееся действие раздражителя.
Информационная роль адаптации состоит в том, что она, отражая своеобразное приспособление рецепторов к стимулам, определяет дискретность раздражения при изменении его интенсивности или местоположения по отношению к рецептору, что также способствует процессам частотной модуляции в рецепторах.
На основании свойств адаптации рецепторов сформулирован закон Вебера— Фехнера.
Закон гласит, что для последующего ощущения раздражение рецепторов должно на некоторую величину превышать ранее действующее раздражение:

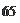 где J— предыдущая сила раздражения; Д/— прирост раздражения.
где J— предыдущая сила раздражения; Д/— прирост раздражения.

