2020-06-12
2020-06-12 219
219Существуют два варианта пространственного картирования ERD/ERS (ERD — event related desynchronization - вызванная десинхронизация, ERS - вызванная синхронизация) На рис в спектрах ЭЭГ человека наблюдается отчетливый мю-ритм как в состоянии с открытыми глазами, так и в состоянии с закрытыми глазами. Данный испытуемый также выполнял двухстимульную GO/NOGO и слуховую oddball-задачи. Для оценки вызванной Десинхронизации в ответ на стимулы, предполагавшие двигательный ответ (нажатие на кнопку), анализ был проведен в соответствии с разработанным G. Pfurtscheller методом. В верхней части рис представлены вызванная десинхронизация/синхронизация между первым предупреждающим стимулом и вторым, который может быть или GO-стимулом (запускающим двигательную реакцию), или NOGO-стимулом (не требующим двигательного ответа). Можно заметить, что подготовительный период сопровождается десинхронизацией мю-ритма в контралатеральном полушарии и синхронизацией в ипсилатеральном полушарии. Внизу на рис. представлены вызванная десинхронизация/синхронизация в ответ на слуховые стимулы: редкий стимул-мишень, предъявление которого требовало нажатия правым пальцем на кнопку, и стимул «не мишень», в ответ на который нажимать на кнопку не требовалось. Можно видеть, что предъявление только стимула-мишени вызывает сильную десинхронизацию мю-ритма.
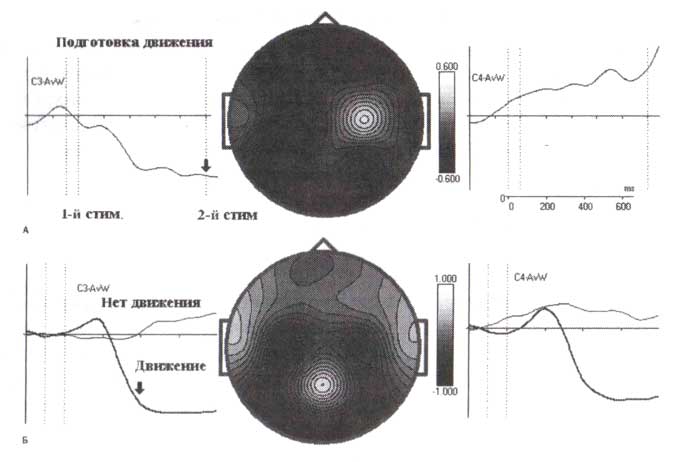
Вызванная десинхронизация во время подготовки к движению и во время самого движения.
А. Вызванная десинхронизация, вычисленная для здорового взрослого субъекта, выполняющего двухстимульную GO/NOGO-задачу. Предъявляются пары изображений животных и растений в различных комбинациях, задача — нажать кнопку в ответ на два предъявленные подряд изображения животных. Обратите внимание на десинхронизацию мю-ритма с левой стороны, сопровождаемую вызванной синхронизацией в правой стороне во время подготовки к движению.
Б. Вызванная десинхронизация, вычисленная для того же самого субъекта, выполняющего слуховую oddball-задачу. Значительная вызванная десинхронизация наблюдается при движениях в ответ на целевой стимул. Обратите внимание, что десинхронизация в одной области сопровождается противоположной реакцией — синхронизацией — в смежной области.
Было показано, что мю-ритмы могут быть дифференцированы на основе их пространственной локализации. Например, области лица и ноги в сенсомоторной коре производят различные мю-ритмы. Мю-ритмы могут разделяться по частотам. Более низкочастотный (8—10 Гц) мю-ритм показывает широко распространенный неспецифический к типу движений паттерн вызванной десинхронизации, сходный для движений пальцев или ступней, тогда как более высокочастотный (10—13 Гц) мю-ритм демонстрирует более сфокусированный, специфический для типов движения паттерн вызванной десинхронизации, отличающийся для движений ступней и пальцев. Оба эти ритма блокируются перед и во время движений, и поэтому их можно рассматривать как мю-ритмы, хотя их реактивные компоненты различны.
Согласно данным, десинхронизация высокочастотного мю-ритма соматотопически специфична и топографически ограничена. Десинхронизация этого ритма начинается в области контралатеральной руки за две секунды перед началом сознательного движения ипсилатеральной руки. Десинхронизация часто сопровождается синхронизацией в центрально-теменных областях, она была названа «очаговой (локальной) десинхронизацией, окруженной зоной синхронизации». В целом этот термин описывает наблюдение, что десинхронизация ритмичной альфа-активности происходит не в изоляции, а может сопровождаться синхронизацией в соседних корковых областях. Области латеральной синхронизации могут соответствовать той же самой сенсорной модальности или другой модальности.
Был предложен таламический селективный, воротный механизм. Та же самая таламическая модель, использовалась для объяснения возникновения сонных веретен, может быть применена для объяснения генерации мю-ритма. Определяющую роль в его схеме играют взаимодействия между таламокорковыми клетками и тормозными нейронами таламического ретикулярного ядра. Последнее формирует топографически организованный тормозной механизм обратной связи, который способен управлять информационным потоком через таламус. Эту функцию управления таламических ретикулярных тормозных нейронов называют «селекцией, отбором, или воротным механизмом». Когда ионные ворота закрыты, таламические нейроны разряжаются в виде пачек спайков, которые проявляются в корковом мю-ритме. Когда ионные ворота открыты, таламические нейроны разряжаются в тоновом режиме, который проявляется в десинхронизации мю-ритма. Согласно предложенному механизму, латеральное торможение, которое существует между ретикулярными нейронами, отвечает за явление «локальной десинхронизации, окруженной зоной синхронизации». Исходя из всего вышесказанного, можно заключить, что роландические мю-ритмы подавляются в ответ на реальные или воображаемые движения.