 2020-06-12
2020-06-12 125
125
Генетический дрейф царствует над нейтральными мутациями (аллелями), отбор – над полезными и вредными. Отбор, повышающий частоту полезных мутаций, называют положительным. Отбор, отбраковывающий вредные мутации, – отрицательным, или очищающим.
Впрочем, все так хорошо и просто только в больших популяциях. В маленьких ситуация сложнее, потому что отбор и дрейф – две главные движущие силы эволюции – начинают конкурировать друг с другом за контроль над слабовредными и слабополезными мутациями.
Посмотрим, как это происходит. Воспользуемся для этого той же моделью с мюмзиками. Изменим только характер мутации, которая привела к появлению аллеля А2. До сих пор мы считали, что мутация была нейтральной. Поскольку она была нейтральной, обладатели аллелей А1 и А2 имели одинаковую плодовитость. Давайте теперь предположим, что мутация была полезной, что она повысила плодовитость мюмзиков на 5 %. Это можно смоделировать так: пусть мюмзики с генотипом А1 рожают по 20 детенышей, а мюмзики с генотипом А2 – по 21. Начальная частота аллеля А2 пусть будет по‑прежнему равна 0,5. Только теперь мы рассмотрим популяции с разной численностью (N).
На рисунке [см. ниже] показано, как будет меняться частота А2 в том случае, если его носители имеют пятипроцентное адаптивное преимущество. Мы видим, что в большой популяции (N = 5000) частота А2 неуклонно растет, приближаясь к единице. Так работает отбор в идеальных для него условиях, т. е. в больших популяциях, где влияние дрейфа на полезные и вредные аллели пренебрежимо мало. Форма у кривой довольно правильная, и это наводит на мысль, что ее можно описать какой‑нибудь математической формулой. Это действительно так, но выводить формулу мы не будем, чтобы не утомить читателей (а любители математики могут сделать это самостоятельно)[19].
Перед нами, между прочим, важнейший эволюционный процесс – аллельное замещение, т. е. вытеснение более приспособленным аллелем менее приспособленного. Процесс идет не слишком быстро. В большой популяции, например состоящей из миллиона особей, для того чтобы зафиксировалась новая полезная мутация, дающая 5‑процентное адаптивное преимущество, требуется около 560 поколений. А ведь 5 % – это серьезное преимущество. Такие мутации – редкость. Ждать, пока зафиксируется мутация с преимуществом в 1 %, придется уже 2800 поколений! Тем не менее в большой популяции отбор «чувствует» даже самую незначительную разницу в приспособленности. Это обеспечивает эффективную (пусть и медленную) фиксацию слабополезных мутаций и элиминацию слабовредных[20].
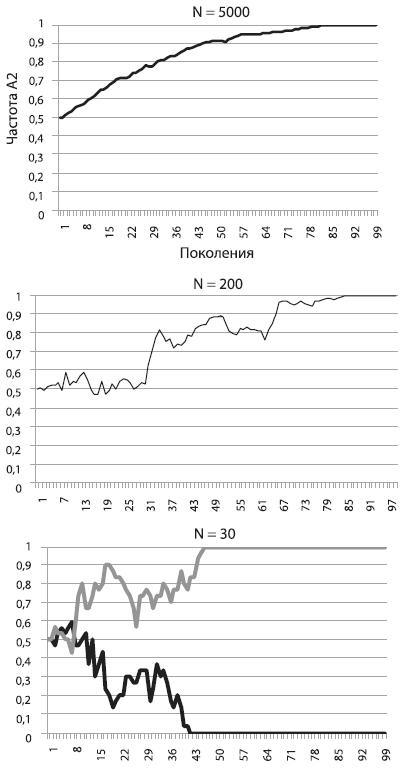
На среднем графике мы видим, какая судьба ждет такую же полезную мутацию, повышающую приспособленность на 5 %, в популяции меньшего размера (N = 200). Частота полезного аллеля и в этом случае росла и в конце концов достигла 1 (мутация зафиксировалась), но путь был труден и извилист. Пожалуй, нашей мутации могло и не повезти, ее шансы на фиксацию не были стопроцентными.
Наконец, на правом графике мы видим, что происходит с точно таким же полезным аллелем в совсем крошечной популяции (N = 30). На рисунке показаны результаты двух запусков модели. В одном случае мутация зафиксировалась, в другом – элиминировалась. Не правда ли, картинка похожа на результат работы дрейфа, а не отбора?
Так оно и есть. В этом заключается главный урок, который мы можем извлечь из наших экспериментов. Чем меньше популяция, тем слабее в ней власть отбора и тем могущественнее дрейф. В маленьких популяциях слабополезные и слабовредные мутации начинают вести себя фактически как нейтральные. Их частоты «случайно блуждают», пока не наткнутся на верхний или нижний порог. В маленьких популяциях, где царствует дрейф, небольшие различия в приспособленности становятся невидимыми для отбора. Поэтому слабовредная мутация запросто может зафиксироваться, а слабополезная – элиминироваться.
Хорошо это или плохо? В большинстве случаев, конечно, плохо. Потеря слабополезных мутаций мешает маленькой популяции приспосабливаться к меняющимся условиям. Бесконтрольное накопление слабовредных мутаций и вовсе может поставить ее на грань вымирания. Именно в этом, между прочим, видят биологи одну из причин того, что крупные животные вымирают в среднем чаще, чем мелкие. У крупных животных, таких как носороги или слоны, популяции не могут быть такими же большими, как у мышей или насекомых. Это снижает приспособляемость крупных животных.
Но у этой медали есть и обратная сторона. У маленьких популяций больше шансов выйти из «ловушки локального оптимума», т. е. сползти с невысокого пика ландшафта приспособленности и взобраться на другой, повыше. Ведь отбор гонит организмы вверх и только вверх. Если популяция велика и малейшее различие в приспособленности «заметно» для отбора, спуск по склонам становится невозможен. Однажды взобравшись на одинокий холм, большая популяция уже никогда с него не слезет. Что касается дрейфа, то он ведет организмы по ландшафту приспособленности хаотическим образом, не замечая подъемов и спусков. Если популяция невелика и дрейф силен, у организмов есть шанс иногда двигаться не только вверх, но и немного вниз (и в сторону). Спустившись в ложбинку, организмы могут «обнаружить», что отсюда есть другой, более перспективный подъем. Если, конечно, не вымрут раньше, чем на него наткнутся.








