 2020-06-12
2020-06-12 118
118
Устойчивость к ядам, вирусам, бактериям и прочим паразитам, защитная окраска, превращение опадающих семян в неопадающие – все это примеры простых адаптаций, для развития которых бывает достаточно одной‑двух удачных мутаций, поддержанных отбором. Более глубокие преобразования складываются из десятков и сотен подобных «мелочей». Одна простая полезная мутация может изменить ландшафт приспособленности для организма (или вывести его в новую область этого ландшафта) – например, изменить взаимоотношения организма со средой, сделав возможной жизнь в условиях, доселе неприемлемых, – и в результате какие‑то другие мутации, прежде бывшие вредными, станут полезными и рано или поздно зафиксируются, открыв возможности для новых изменений.
Поскольку для крупных эволюционных преобразований требуется последовательное закрепление множества мутаций, такие изменения трудно «расшифровать» на молекулярном уровне, а уж пронаблюдать воочию и вовсе невозможно из‑за длительности процесса. Те случаи, которые все‑таки удается расшифровать, – это, по‑видимому, редкие, нетипичные случаи, когда крупное изменение обеспечивается всего несколькими мутациями. Но такие случаи есть, и мы пока не знаем наверняка, так ли уж мала их роль на больших – эволюционно значимых – отрезках времени. Мы познакомимся с одним из таких случаев, когда всего две мутации обеспечили интересное и важное адаптивное приобретение у многоклеточного животного – нематоды (круглого червя). В результате двух мутаций нематоды приобрели способность к самооплодотворению – стали гермафродитами. Этот пример важен нам также для того, чтобы показать, как в ходе эволюции подстраиваются друг к другу разные гены, регулирующие определенную функцию.
У большинства нематод, как и у многих других животных, пол определяется генетически, при помощи половых хромосом. Если в оплодотворенном яйце две X‑хромосомы, из яйца разовьется самка, если одна – самец (Haag, 2005). Однако у двух наиболее изученных видов нематод, Caenorhabditis elegans и C. briggsae, особи с двумя X‑хромосомами – не самки, а гермафродиты. Их гонады (половые железы) на поздних личиночных стадиях производят спермии. Нематоды утратили в ходе эволюции жгутики, поэтому и спермии у них безжгутиковые. Они похожи на амеб и передвигаются при помощи псевдоподий. У гермафродитов безжгутиковые спермии поздних личинок переползают в специальные хранилища – сперматеки – и здесь ждут своего часа. Гонады взрослых гермафродитов производят уже не спермии, а яйцеклетки. Они могут быть оплодотворены как собственными спермиями из сперматеки, так и спермиями самца в результате спаривания.
Предки C. elegans и C. briggsae были нормальными раздельнополыми червями. Это следует, в частности, из того, что все прочие виды рода Caenorhabditis – раздельнополые. По‑видимому, гермафродитизм у C. elegans и C. briggsae является «эволюционно молодым», новым признаком. Чтобы разобраться, как он возник, понадобились выдумка, знание геномов нескольких видов нематод и эксперименты, осуществленные американскими биологами (Baldi et al., 2009). Вот как это было.
Для начала уточним, какие гены работают при формировании половых различий у нематод. Наличие одной (а не двух) X‑хромосом у самцов служит триггером, включающим синтез белка HER‑1, который ингибирует белок TRA‑2. Подавление активности TRA‑2 через пару промежуточных шагов приводит к уничтожению белка TRA‑1, функция которого состоит в том, чтобы отключать ряд ключевых генов, направляющих развитие по «мужскому» пути (в том числе ген fog‑3, контролирующий сперматогенез)[29]. У гермафродитов белок HER‑1 не образуется, но они все равно производят спермии, потому что активность гена tra‑2 на стадии личинки подавляется белками FOG‑2 и GLD‑1. Таким образом, гермафродиты получаются из‑за введения в игру дополнительного правила, связанного с регуляцией гена tra‑2.
Запутались? Ничего страшного, это в порядке вещей. Регуляторные сети, управляющие развитием животных, как правило, сложны, громоздки и неоптимальны[30]. К счастью, их не нужно учить наизусть никому, кроме студентов соответствующих специальностей. Чтобы облегчить дело, можно представить устрашающее описание из предыдущего абзаца в виде графической схемы (см. следующую страницу).
На таких схемах стрелочками обозначают положительные влияния (включение гена, активация белка), а «тупичками» – отрицательные (ингибирование белка, подавление активности гена).
Так вот, зная все это, исследователи подумали, что можно попробовать превратить самок раздельнополого вида C. remanei в гермафродитов, подавив активность какого‑нибудь гена, блокирующего сперматогенез. Например, гена tra‑2.
Начали они с того, что отключили tra‑2 при помощи РНК‑интерференции[31]. В результате из яиц с двумя X‑хромосомами развились вместо самок обыкновенные самцы, производящие нормальные спермии, но никаких яйцеклеток.
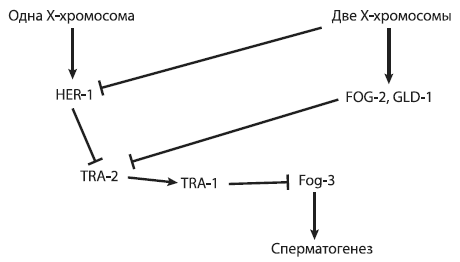
Схема регуляции развития пола у нематоды C. elegans, у которой вместо самок гермафродиты.
Тогда ученые справедливо рассудили, что они, видимо, перестарались. Ведь у настоящих гермафродитов активность гена tra‑2 хоть и снижена, но не до нуля. Когда эту ситуацию удалось воспроизвести у C. remanei, уменьшив экспрессию tra‑2, на свет появились животные, которых авторы назвали псевдогермафродитами. Тело у них было «женское», но в гонадах формировались одновременно и яйцеклетки, и спермии. Правда, эти существа оказались неспособны к самооплодотворению (отсюда и приставка «псевдо»). После скрещивания с самцом превдогермафродиты откладывали оплодотворенные яйца, хоть и в меньшем количестве, чем обычные самки. Это значит, что яйцеклетки у них более или менее нормальные, а неспособность к самооплодотворению, скорее всего, объясняется дефектами спермиев.
Действительно, спермии псевдогермафродитов оказались нормальны по всем параметрам, кроме одного: они неактивны, никуда не ползут, и в частности не перебираются в сперматеки.
У обычных нематод спермии активируются (начинают ползать) под воздействием белков, содержащихся в семенной жидкости. Сохранили ли спермии псевдогермафродитов способность активироваться под действием этих белков? Чтобы это проверить, авторы скрещивали псевдогермафродитов с самцами C. elegans. Самцы C. elegans охотно спариваются с самками C. remanei, но эти браки бесплодны (как‑никак два вида разошлись, судя по показаниям молекулярных часов, около 80 млн лет назад, а для развития генетической несовместимости обычно хватает нескольких миллионов лет[32]). Идея такого спаривания как раз и состояла в том, чтобы половые клетки от разных видов не сливались, но при этом псевдогермафродиты могли получить нормальные белки семенной жидкости.
И когда после этого адюльтера псевдогермафродиты C. remanei отложили оплодотворенные яйца, некоторые из них оказались жизнеспособными! Из них вывелись нормальные самки C. remanei (не псевдогермафродиты, потому что им никто не подавлял активность гена tra‑2). Это означает, что спермии у псевдогермафродитов получились нормальными, не хватает только активирующих белков.
Это открытие сузило круг потенциальных генов‑мишеней, на которые нужно воздействовать, чтобы превратить псевдогермафродитов в полноценных гермафродитов. «Подходящим» геном оказался swm‑1, который кодирует белок, подавляющий активность других белков – протеаз, активирующих спермии. Ген swm‑1 отвечает за предотвращение преждевременной активации спермиев у самцов C. elegans, но у него есть и другие функции. Ученые понизили активность этого гена у псевдогермафродитов – и те, к большой радости исследователей, приобрели способность к самооплодотворению.
Таким образом, для появления нового признака – гермафродитизма – достаточно уменьшить активность двух генов, входящих в два разных регуляторных каскада. Для этого нужны две мутации. Изменение активности того или иного гена в результате случайной мутации – дело самое обычное. Проблема в другом: одновременное возникновение двух мутаций, полезных только вместе, но не по отдельности, крайне маловероятно. Могли ли они появиться последовательно или это равносильно преодолению пропасти в два прыжка? Исследователи рассмотрели два возможных сценария.
1. Сначала произошла мутация, уменьшившая активность swm‑1 у самок, что позволяет активировать собственные спермии (если они есть). Эта мутация поначалу была нейтральной, потому что самки еще не умели производить спермии. В дальнейшем возникла мутация, понизившая активность tra‑2. Эта мутация сразу оказалась полезной (т. е. была поддержана отбором), потому что превратила самок в самодостаточных гермафродитов (о полезности гермафродитизма см. ниже). Но могла ли первая мутация не принести вреда, если известно, что ген swm‑1 многофункционален? И вот тут‑то на помощь приходит знание геномов изучаемых червей (недаром в последние годы биологи столько сил и средств тратят на прочтение геномов всевозможных тварей). Оказывается, в геномах раздельнополых видов есть только одна копия swm‑1, а у гермафродитных есть еще и вторая, немного отличающаяся копия. По‑видимому, становлению гермафродитизма способствовала дупликация (удвоение) этого гена, что и позволило снять «адаптивный конфликт». Одна из копий изменилась, чтобы обеспечить активацию спермиев у гермафродитов, а вторая продолжила выполнение остальных функций[33].
2. Второй сценарий начинается с мутации, снизившей активность tra‑2. Это привело к появлению псевдогермафродитов. Лишь после этого возникла и закрепилась мутация, снижающая активность swm‑1. Однако псевдогермафродиты фактически являются всего лишь неполноценными самками: к самооплодотворению они не способны, а с ролью самок справляются хуже, чем настоящие самки. Поэтому на первый взгляд кажется, что отбор должен был отсеять первую мутацию. Но вспомним, что самки C. remanei привлекательны для самцов других видов, а спаривание с этими самцами дает им возможность самооплодотвориться. Эта особенность могла стать «мостиком», с помощью которого эволюционирующий вид сумел преодолеть опасный промежуточный этап и дождаться возникновения второй мутации. В некоторых ситуациях псевдогермафродиты, способные к самооплодотворению после спаривания с самцами других видов, могли иметь адаптивное преимущество – например, когда численность популяции критически снижалась.
Таким образом, при ближайшем рассмотрении оказывается, что червям не нужно было «преодолевать пропасть в два прыжка». Там все‑таки были мостики.
Изменение активности гена в сторону уменьшения или увеличения может произойти в результате самых разных мутаций. Это могли быть мутации в регуляторных областях самих генов tra‑2 и swm‑1, или мутации генов‑регуляторов, управляющих их работой, или мутации регуляторов регуляторов, и т. д. Важно, что в такой ситуации – когда оказывается выгодно уменьшить или увеличить экспрессию какого‑либо гена – вероятность того, что случайная мутация, меняющая активность гена, окажется полезной, а не вредной, приближается к 1/2, т. е. становится чрезвычайно высокой. Скорее всего, именно поэтому очень многие «прогрессивные» эволюционные преобразования, как теперь выясняется, были связаны с изменениями уровня активности генов, а не их белок‑кодирующих последовательностей. Между прочим, это относится и к эволюции человека (Gilad et al., 2006).
Появление новых признаков путем изменения активности генов – один из магистральных путей эволюции. Почему? Да прежде всего потому, что это просто. В инструкциях по устранению неисправностей в работе электроприборов неизменно присутствует «мудрый совет», раздражающий многих: проверьте, включена ли вилка в розетку. За ним кроется универсальный принцип: во многих случаях нужного эффекта можно добиться, манипулируя только выключателями и не развинчивая весь механизм.
Конкретные мутации, ответственные за снижение активности генов tra‑2 и swm‑1 у предков C. elegans и C. briggsae, возможно, были разными. Например, известно, что у гермафродитов первого вида в подавлении активности tra‑2 участвует ген fog‑2, отсутствующий у второго вида. Предки C. briggsae утратили ген fog‑2, вероятно за ненадобностью. У этого вида ключевую роль в обеспечении нормального сперматогенеза у гермафродитов играет другой ген – she‑1. Этот пример показывает, что в ходе эволюции «переключатели» генетических регуляторных каскадов могут меняться, в то время как структура и функции каскадов остаются прежними.
Это относится и к механизмам определения пола. Разделение на самцов и самок есть у большинства животных. Соответственно, есть и генетические «программы» (большие и сложные) развития по мужскому и женскому пути. Однако переключатели, направляющие развитие по одному из двух путей, часто меняются в ходе эволюции. Например, у тех же нематод многие виды перешли от хромосомной детерминации пола к «средовой», т. е. пол у них зависит не от генов, а от условий, в которых проходит развитие. Мы теперь понимаем, что такие эволюционные изменения относятся к числу высоковероятных. Точно так же вы можете заменить кнопку электрического выключателя в своей комнате на систему последовательно и параллельно соединенных тумблеров, реле или повесить шнурок с кисточкой, не меняя при этом проводку и люстру. Управлять освещенностью, возможно, станет удобнее, но сам «фенотип» от этого не изменится: свет по‑прежнему будет или включен, или выключен.
Ну и последнее: зачем вообще кому‑то понадобилось переходить к гермафродитизму? Как могли мутации, превратившие самок в гермафродитов, оказаться полезными для предков C. elegans и C. briggsae? Дело в том, что в некоторых ситуациях гермафродитизм дает очевидное преимущество[34]. Например, если участки, пригодные для жизни нематод данного вида, встречаются очень редко (это вполне справедливо для представителей рода Caenorhabditis) и вероятность того, что хотя бы один червь попадет на новый, незаселенный участок, очень мала, то вероятность того, что на этот участок попадут сразу два червя – самец и самка, – будет и вовсе ничтожной. Естественно, в такой ситуации преимущество получат черви, способные размножаться без посторонней помощи, путем самооплодотворения. Или, на худой конец, с помощью самцов других видов.
Гены взаимной дружбы
В книге «Рождение сложности» немало говорилось об эволюционной роли симбиоза. На одних мутациях и отборе можно уехать далеко, но возможность комбинировать эволюционные «достижения» разных организмов в одном симбиотическом сверхорганизме открывает еще более впечатляющие перспективы. Впрочем, чтобы наладить с кем‑то эффективный симбиоз, без мутаций и отбора тоже не обойтись. Такие договоры на гербовой бумаге не подписываются, тут нужно тщательно приладиться друг к другу.
Одним из таких межорганизменных «договоров» является симбиоз наземных растений с почвенными грибами и бактериями. Эволюционный успех наземных растений во многом был обеспечен именно этими взаимовыгодными отношениями: грибы и бактерии снабжают растение соединениями азота и фосфора, получая взамен углеводы, образуемые растением в ходе фотосинтеза. Ведь сами наземные растения так и не научились фиксировать азот.
Самой древней разновидностью такого симбиоза является микориза, известная в двух вариантах: более простая эктомикориза (гриб не проникает внутрь растительных клеток) и эндомикориза, или арбускулярная микориза, при которой гифы гриба врастают внутрь клеток корня. Судя по палеонтологическим данным, арбускулярная микориза существовала уже у древнейших наземных растений, робко пытавшихся освоить негостеприимную сушу 450 млн лет назад, в ордовикском периоде. Весьма вероятно, что без симбиоза с грибами эти попытки были бы обречены на провал – или, что еще вероятнее, их бы не было вовсе. Некоторые данные указывают на то, что симбиоз с грибами зародился у зеленых водорослей, предков наземных растений, еще в водной среде, что и обеспечило возможность освоения суши. О древности арбускулярной микоризы свидетельствует, помимо прочего, ее широкое распространение во всех группах наземных растений.
Помимо грибной микоризы пользуется спросом и симбиоз с азотфиксирующими бактериями. Отношения растений с азотфиксаторами тоже имеют древнюю историю, однако в большинстве случаев речь идет о внеклеточных бактериальных симбионтах. Только некоторые покрытосеменные (цветковые) растения сравнительно недавно научились культивировать бактерии внутри клеток своих корней, в особых органах – клубеньках. Клубеньковые симбиозы бывают двух типов: 1) симбиоз бобовых с ризобиями (бактериями из группы альфапротеобактерий); 2) актинориза – симбиоз с актинобактериями рода Frankia. Актинобактерии образуют многоклеточный мицелий наподобие грибов; раньше их относили к грибам и называли актиномицетами.
Клубеньковые симбиозы встречаются только в четырех группах (порядках) покрытосеменных: у бобовых (Fabales), розовых (Rosales), тыквенных (Cucurbitales) и буковых (Fagales), причем не у всех, а только у части представителей. Недавно на основе молекулярно‑генетических данных было установлено, что эти четыре порядка представляют собой монофилетическую кладу, т. е. группу, происходящую от общего предка и включающую всех его ныне здравствующих потомков. Вероятно, у общего предка этой группы произошли какие‑то генетические изменения, обусловившие возможность развития клубенькового симбиоза. Одни представители группы впоследствии воспользовались этой возможностью, другие нет.
Симбиоз бобовых с ризобиями изучен лучше, чем актинориза. Но у двух типов клубеньковых симбиозов обнаружилось много общего: по меньшей мере семь генов задействованы в обоих симбиозах. Эти гены получили название «общих генов симбиоза». По‑видимому, при становлении клубенькового симбиоза была использована древняя генетическая программа, сложившаяся изначально для обслуживания внутриклеточного симбиоза с грибами. Для полноты картины хотелось бы уяснить, что же изменилось в генах при становлении тесных отношений с азотфиксаторами.
В 2008 году британские и германские биологи (Markmann et al., 2008) взяли разные группы цветковых и проанализировали строение белков, кодируемых «общими генами симбиоза». Оказалось, что у всех цветковых большинство этих белков имеют почти одинаковую, «консервативную» структуру. И только один из них оказался вариабельным. Белок этот называется SYMRK (symbiosis receptor kinase). Он различается у клубеньковых растений, двудольных и однодольных.
Строение SYMRK, а точнее строение его доменной части (домен – функциональная часть или блок белковой молекулы, содержащий узнаваемый аминокислотный «мотив»), говорит кое‑что о специализации этого белка в клетке. У него есть, во‑первых, трансмембранный домен, который, как видно из названия, располагается в толще клеточной мембраны (для белка клеточная мембрана и вправду толстая и вместительная). Во‑вторых, внутриклеточная часть белка содержит домен протеинкиназы, функция которого состоит в переносе фосфата с АТФ на какой‑нибудь белок. Этот процесс называется фосфорилированием. Таким способом многие рецепторные белки передают полученный извне сигнал внутрь клетки, поскольку фосфорилирование белков меняет их свойства – например, переводит их в активное состояние из неактивного. И трансмембранный домен, и домен протеинкиназы в белке SYMRK почти одинаковы у всех цветковых. Различия сосредоточены во внеклеточной части белка, которая выполняет рецепторную функцию, т. е. улавливает внешний химический сигнал. Исследователи выявили три варианта этой вариабельной внеклеточной части белка SYMRK:
1) «длинный вариант», характерный для растений, образующих клубеньки, и их близких родственников;
2) «средний вариант», характерный для дальних родственников клубеньковых растений;
3) «короткий вариант», характерный для однодольных.
Грибная арбускулярная микориза встречается у обладателей всех трех вариантов гена. Клубеньки любого типа (содержащие ризобии или актинобактерии) встречаются только у обладателей «длинного» варианта гена SYMRK. Очевидно, желающие приобрести клубеньки позаботились о своевременном распознавании дефицитных симбионтов. Длина нуклеотидной последовательности увеличивается за счет включения в ключевую область двух фрагментов (доменных мотивов) других генов. Оба эти фрагмента были не изобретены заново, а «заимствованы» у генов белков, имеющихся в геноме растений.
Очевидно, приобретение «длинного» варианта SYMRK как раз и было тем ключевым событием, которое создало предпосылки для развития клубеньковых симбиозов – причем «генетическая программа» клубенькового симбиоза представляет собой модификацию «генетической программы» арбускулярной микоризы. Это проверили в серии экспериментов.
В первом эксперименте у растения датиска (Datisca glomerata), корни которого в норме образуют арбускулярную микоризу и актиноризу, отключили ген SYMRK. В результате растение утратило способность к формированию обоих симбиозов – и с грибом, и с актинобактерией Frankia. Значит, ген SYMRK необходим и для грибного, и для бактериального симбиоза, у них единая генетическая основа.
Второй эксперимент показал, что ген SYMRK не служит для распознавания конкретных бактерий‑симбионтов. Бобовое растение лядвенец японский (Lotus japonicus) образует клубеньки с бактерией‑ризобией Mesorhizobium loti, а люцерна (Medicago truncatula) – с бактерией Sinorhizobium melioti. Мутантной люцерне с испорченным геном SYMRK, не способной формировать никакой симбиоз вообще, пересадили ген SYMRK от лядвенца. Эта операция полностью восстановила способность люцерны образовывать симбиоз. При этом трансгенная люцерна стала образовывать клубеньки со «своей» исконной бактерией Sinorhizobium, а вовсе не с Mesorhizobium. Также мутантному лядвенцу пересаживали ген SYMRK от других бобовых и их бесклубеньковых родственников, например настурции, и во всех случаях его пошатнувшиеся отношения со своим Mesorhizobium восстанавливались. Таким образом, SYMRK отвечает не за узнавание и выбор симбионта, а только за общую способность формировать внутриклеточный симбиоз с бактериями. Узнавание осуществляется другими белками, какими именно – пока не установлено.
В третьем эксперименте снова использовали мутантную форму лядвенца японского, не образующую ни арбускулярной микоризы, ни клубеньков. Растениям пересаживали «средний» вариант гена, взятый у помидора, и «короткий» вариант, позаимствованный у риса. В обоих случаях у мутантного лядвенца восстановилась способность к формированию арбускулярной микоризы, но не клубеньков. Следовательно, укороченные варианты гена SYMRK достаточны для грибной микоризы, но не для клубеньковых симбиозов.
Белок SYMRK необходим для формирования особых внутриклеточных структур – симбионтоприемников или «преинфекционных нитей», которые впоследствии заселяются симбиотическими бактериями (и тогда их уже называют «инфекционными нитями»). Похожие структуры образуются в клетках корней и для принятия грибных симбионтов (в случае арбускулярной микоризы). Сходство в строении этих симбионтоприемников отражает единство генетической программы, отвечающей за формирование всех трех типов внутриклеточного симбиоза: арбускулярной микоризы, симбиоза с ризобиями и актиноризы.
Полученные результаты подтверждают гипотезу, согласно которой способность к формированию клубеньковых симбиозов развилась на основе древней генетической программы арбускулярной микоризы. Ключевое эволюционное событие заключалось в том, что клетки корней приобрели способность реагировать формированием «симбионтоприемников» не только на присутствие симбиотических грибов, но и на близость азотфиксирующих бактерий. А если пристальней вглядеться в молекулярные подробности этого события, то увидим, что для этого понадобилось перенастроить рецепторную часть одного мембранного белка (SYMRK). Этот белок пристроил в свое пользование два дополнительных участка из других имеющихся под рукой генов. Можно сказать, что для налаживания симбиоза организмов использовался своеобразный молекулярный, генный симбиоз. Принципы конструирования нового схожи и на уровне молекул, и на уровне органов, организмов и популяций.
Возникновение клубеньковых симбиозов – исключительное по своей полезности приобретение, имеющее к тому же важное биосферное значение. Казалось бы, для такого «революционного» нововведения потребуется масса приспособлений, реорганизация больших областей генома. Но нет, понадобилось всего лишь научиться узнавать во внешней среде новый объект, а для этого – чуть‑чуть видоизменить белок‑рецептор. Когда речь идет о внешне сложной проблеме, помогает внимательное разглядывание подробностей (как в примере с белоногими хомячками, чья светлая защитная окраска складывается всего лишь из расширения светлой полосы на шерстинках): ключевое изменение зачастую оказывается простым и легкодостижимым.
–––––
Между микро‑ и макроэволюцией нет принципиальной разницы
Из истории азотфиксирующих симбиозов можно извлечь важный урок. Мы увидели, что важное новшество, такое как появление клубеньковых симбиозов – событие без преувеличения глобальной, биосферной значимости! – реализовалось в ходе эволюции по той же схеме, что и приобретение малярийным плазмодием устойчивости к хлорохину. В обоих случаях ключом к приобретению нового признака стало изменение активного центра белка, отвечающего за избирательное связывание тех или иных веществ. Правда, плазмодий обошелся нуклеотидной заменой, а у растений произошла внутригеномная рекомбинация – перетасовка участков генов. Но это, по правде сказать, пустяк. Гены могли перетасоваться и у плазмодия (с примерами таких событий мы встретимся в следующих главах).
Подобные параллели между самыми крупными и самыми ничтожными эволюционными событиями убеждают биологов в том, что между так называемыми микроэволюцией и макроэволюцией[35] нет принципиальной разницы. Это исключительно вопрос масштаба. Посмотрите на эволюцию в лупу – увидите микроизменения. Взгляните издалека – увидите более значительные макроперемены. Но в основе тех и других лежат сходные механизмы.
На единую природу микро‑ и макроэволюции указывают и другие факты, в том числе фундаментальное сходство, если не сказать тождество, внутри‑ и межвидовой изменчивости. Если мы начнем сравнивать генетические различия между двумя особями одного вида с различиями, существующими между особями разных видов, то увидим в буквальном смысле одно и то же: замены нуклеотидов («однонуклеотидные полиморфизмы»), вставки и выпадения нуклеотидов («инделы»), транспозиции (перемещения фрагментов ДНК из одного места генома в другое), инверсии (повороты фрагментов ДНК на 180°), вариации по числу копий повторяющихся фрагментов и т. д. Крупные хромосомные перестройки, такие как слияние двух хромосом в одну или, наоборот, разделение, в пределах одного вида встречаются реже, чем при межвидовых сравнениях, но все же встречаются. Такие перестройки чреваты снижением плодовитости гибридного потомства (т. е. потомства от скрещивания родительских особей с разным числом хромосом), но не являются непреодолимым барьером для гибридизации, как показывают многочисленные примеры видов, в пределах которых число хромосом варьирует[36]. Например, среди диких кабанов, проживающих в Испании, встречаются особи с 36, 37 и 38 хромосомами (Nombela et al., 1990).
Биологи не сразу пришли к пониманию тождества внутри‑ и межвидовой изменчивости. Довольно долго допускалось существование особых «макроизменений», которые приводят к появлению новых видов и которые принципиально отличаются от внутривидовой изменчивости. Стремительное развитие методов изучения ДНК в последние два десятилетия позволило проверить эти предположения. Проверки они не выдержали. На сегодняшний день о них можно смело забыть.
Важно, что внутри‑ и межвидовые различия идентичны не только на качественном уровне, но и на уровне количественных соотношений. К примеру, если мы сопоставим генетические различия, имеющиеся между разными людьми, с теми различиями, которые отделяют нас от шимпанзе, то увидим, что эти две группы различий одинаковы по многим статистическим параметрам: по соотношению значимых и незначимых замен в белок‑кодирующих генах, по соотношению однонуклеотидных замен и выпадений или вставок нуклеотидов в некодирующих областях и т. д. Между двумя людьми, конечно, различий меньше, чем между человеком и шимпанзе (примерно в 10–20 раз), но различия эти – одной и той же природы.
Например, недавно международная команда генетиков опубликовала результаты сравнения геномов 29 видов плацентарных млекопитающих (Lindblad‑Toh et al., 2011). Ученые выявили 3,6 млн функциональных участков ДНК, находящихся под действием очищающего отбора. Эти участки, мутации в которых не являются нейтральными, составляют примерно 5,5 % генома у плацентарных. Около трети из них соответствуют белок‑кодирующим, остальные две трети – регуляторным последовательностям. Выявлено 280 тыс. регуляторных участков, происходящих из фрагментов мобильных генетических элементов; 563 участка, эволюция которых шла ускоренными темпами у предков человека после их отделения от предков шимпанзе. Но для нас сейчас важно другое. Когда авторы сопоставили полученные данные с имеющейся информацией по внутривидовой генетической вариабельности Homo sapiens, оказалось, что эти массивы данных прекрасно согласуются друг с другом. Те участки генома, которые мало отличаются у разных видов плацентарных (т. е. являются консервативными – медленно меняющимися в ходе эволюции), в пределах человеческой популяции тоже имеют низкую вариабельность. И наоборот: те участки, которые у разных людей могут сильно отличаться друг от друга, у других плацентарных тоже изменчивы. Более того, многие сайты (нуклеотидные позиции) с ограниченной эволюционной пластичностью (например, позиции, в которых может стоять нуклеотид Г или Т, но не А и не Ц), варьируют одинаковым образом как внутри человеческой популяции, так и у разных видов плацентарных. Это значит, что очищающий отбор, действовавший на геномы различных плацентарных, продолжал схожим образом действовать и на геномы ближайших предков современного человечества – а потому и изменчивость, накопление которой определяется характером очищающего отбора, оказывается сходной внутри вида и между видами.
Итак, изменчивость, определяющая различия между особями одного вида, – это, по сути, та же самая изменчивость, что определяет различия между видами, родами, семействами и т. д. Дайте ей только время, чтобы накопиться.
Эти факты говорят о единстве микро‑ и макроэволюции, т. е. внутри‑ и межвидовых эволюционных изменений. Пользуясь модным ныне словечком, можно сказать, что эволюция фрактальна: большое отражается в малом, малое – в большом. Поэтому изучение мельчайших, только в микроскоп заметных событий (а нам их изучать легче всего, учитывая скоротечность жизни) дает адекватное представление и о событиях гораздо большего масштаба.
–––––
Глава 3
Секс
Кроме размножения (в основе которого лежит репликация ДНК), наследственности (основанной на специфическом спаривании нуклеотидов), мутаций (включая перетасовку фрагментов ДНК внутри генома) и естественного отбора (влияния мутаций на эффективность размножения) у земной жизни есть еще одна важная шестеренка, еще один незаменимый механизм создания нового, без которого эволюция едва ли смогла бы даже стартовать, не говоря уж о сотворении китов и зябликов.
Биологи, особенно англоязычные, часто называют явление, о котором идет речь, простым и выразительным словом «секс». В русском языке «секс» имеет иной спектр смысловых оттенков, что чревато путаницей. Поэтому давайте уточним, что именно мы будем иметь в виду под сексом. Мы будем иметь в виду любые способы комбинирования в одном геноме фрагментов разных геномов. Это можно еще назвать «межорганизменной рекомбинацией», но «секс» короче и яснее. Под такое определение подходит и привычное нам половое размножение, характерное для сложных организмов, и горизонтальный перенос генов, характерный для бактерий, и даже обмен участками между двумя вирусными геномами, заразившими одну и ту же клетку.
Как секс встраивается в классическую триаду эволюции: изменчивость – наследственность – отбор? Это, если подумать, нечто отличное и от первого, и от второго, и от третьего. Это отдельный фактор, который устанавливает новые отношения между наследственностью и изменчивостью. Он существенно увеличивает пользу от изменчивости, позволяя эволюции работать не с целыми геномами, а с отдельными генами и не с индивидуальными организмами, а с популяционными генофондами. Это резко повышает эффективность отбора. Именно секс превращает биологическое разнообразие на всех его уровнях в мощную адаптивную силу. Но вместе с тем применение этой силы заставляет организмы вырабатывать множество приспособлений, постоянно учитывать именно этот четвертый фактор. Вот уж где сексуальная революция формирует совершенно новые отношения и выдвигает жесткие и специфические требования к приверженцам этих новых отношений. В этой главе мы покажем, для чего нужен и как работает четвертый эволюционный фактор – секс.
–––––








