 2020-06-12
2020-06-12 146
146
До какого‑то момента процесс видообразования остается обратимым. Изменение условий может привести к тому, что наметившиеся различия сгладятся, а начавшие расходиться популяции снова объединятся. Это может произойти даже с «хорошими» видами, по поводу которых у специалистов нет сомнений, виды это или все‑таки разновидности. В наши дни основной причиной таких «обратных эволюционных ходов» является антропогенное воздействие на среду. Например, по словам ихтиолога М. В. Мины из Института биологии развития РАН, под угрозой слияния оказались многие виды цихлид озера Виктория (Мина, 2001). Причина – в неразумном решении вселить в озеро нильского окуня, крупного хищника, который быстро размножился в озере и поставил многие местные виды под угрозу физического уничтожения. Что касается видов, еще не съеденных окунем, то некоторые из них начали скрещиваться друг с другом, «растворяя» свои уникальные особенности в смешанных генофондах. Дело в том, что нильский окунь – рыба крупная. Его нельзя просто высушить на солнышке, как испокон веков поступали местные рыбаки с мелкой рыбешкой (теми самыми цихлидами). Окуня нужно коптить, а для этого нужны дрова. Вырубка деревьев по берегам озера усилила эрозию почв. Дожди стали смывать в озеро больше глины и песка. Озерная вода, и без того мутноватая, замутилась еще больше. В мутной воде многие цихлиды перестали различать нюансы окраски потенциальных партнеров, служившие изолирующим барьером между видами. Началась гибридизация.
Пассивное накопление генетических различий по модели Добжанского – Мёллера у цихлид требует обычно от 4 до 14 млн лет для развития полной несовместимости. Возраст озера Виктория намного меньше, поэтому большинство местных цихлид еще сохранили способность к гибридизации. Таким образом, вселение одного‑единственного вида поставило под угрозу «великий эволюционный эксперимент» природы.
Еще один пример обратного хода эволюции исследовали экологи из Швейцарии и Канады. Ученые воспользовалась превосходными данными по сигам из швейцарских горных озер. В их распоряжении имелись профессионально собранные данные 1940‑х годов, а также современные материалы, отражающие разнообразие сигов в начале XXI века (Vonlanthen et al., 2012). В науке так всегда бывает: чем надежнее данные, тем интереснее результат. Не стало исключением и это исследование. Обычно при анализе эволюции оценивают степень расхождения видов, здесь же наблюдали обратное. Ученые зарегистрировали эволюцию, повернувшую вспять.
Использовались данные по 17 озерам в предгорьях Швейцарских Альп. Это глубокие водоемы с хорошо аэрированной водой, изначально олиготрофные, т. е. с малым содержанием органики. В озерах издавна обитали сиги (род Coregonus). Они заселились после освобождения этой территории от ледника (10–12 тыс. лет назад) и начали осваивать новое экологическое пространство. В ходе специализации и разделения ниш в каждом озере образовались «букеты» симпатрических видов (1–5 в каждом озере), в том числе характерные пары экологических аналогов. В каждой такой паре один вид предпочитает мелководья, питается мелкой добычей, нерестует в зимние месяцы; обычно это крупные рыбы с небольшим числом тычинок на жаберных дугах. Второй вид – его контрагент; обычно это мелкая рыба с большим числом жаберных тычинок, предпочитает жить на глубине, а нереститься летом, питается более крупным зоопланктоном. Число жаберных тычинок отражает пищевую специализацию, четко наследуется и считается хорошим маркером видовой принадлежности. Учитывая разночтения систематики рода Coregonus, в озерах насчитывается всего около 25 видов и подвидов сигов.

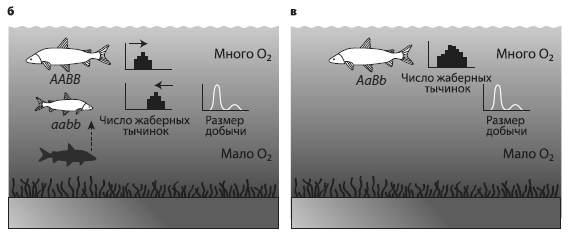
Уменьшение разнообразия сигов в озерах предгорьев Альп. а – ситуация в начале xx века: существует два вида – крупный и мелкий. Крупный обитает в неглубоких водах, мелкий – где поглубже, число жаберных тычинок у крупного меньше (маленькие гистограммы напротив рыбок) в соответствии с размером предпочитаемой добычи (маленькие графики). Кислорода в воде достаточно. б – ситуация после эвтрофикации, когда в придонных слоях воды мало кислорода. Число жаберных тычинок и размер предпочтительной добычи у мелких рыб уменьшается. в – исчезновение глубоководного вида. У мелководного при этом увеличивается генетический полиморфизм и вариабельность по числу жаберных тычинок и размеру добычи. Из McKinnon, Taylor, 2012.
Таково было состояние озерных систем до 1950‑х годов, описанное в классических трудах европейских ихтиологов. Каковы же современные данные? А они таковы, что около четверти видов сигов исчезло из видового реестра, причем вымерли в основном мелкие глубоководные виды. Связано это с загрязнением озер и резким повышением содержания органики в воде. Механизм вымирания на первый взгляд прост. Увеличение количества органики вызывает бурный рост микрофлоры, которая окисляет органику растворенным в воде кислородом. В результате содержание кислорода, особенно в придонных слоях, падает, нерестилища глубоководных рыб оказываются непригодными, молодь гибнет и вид исчезает.
Схема как будто проста, но, если поразмышлять о деталях процесса, все окажется интереснее. Вид быстро исчезает… Совсем исчезает? Ведь исчезнуть можно по‑разному. В данном случае глубоководный вид слился с мелководным. Два вида начали скрещиваться, и в результате уцелевший мелководный вид приобрел некоторые черты исчезнувшего глубоководного. Это было доказано с помощью анализа морфологической (по числу жаберных тычинок) и генетической вариабельности.
В одном случае процесс обратного слияния видов удалось «поймать за хвост», увидеть его в действии. Этот пример исследовали на материале Цюрихского и Валенштадтского озер. Пример хорош тем, что к истории видов прилагается в качестве бонуса еще и история самих озер. Два озера когда‑то были одним целым. В это послеледниковое праозеро заселился один предковый вид. Там же он разделился на два эндемичных вида – мелководный C. duplex и глубоководный C. heglingus. Затем в праозере обособились две части. Оба вида оказались в обоих частях озера – теперь в двух отдельных озерах, и генетический анализ действительно доказывает такое их происхождение. Однако генетическая удаленность видов друг от друга оказалась разной в двух озерах. В Цюрихском озере, сильно эвтрофицированном, они генетически ближе друг к другу и явно скрещиваются, а в чистом Валенштадском озере их генетическая вариабельность не перекрывается. Это значит, что в Цюрихском озере вовсю идет процесс стирания границ между видами: глубоководный вид сливается с мелководным.
Не менее интересен пример с Боденским озером. Там еще осталось три вида сигов, в том числе глубоководный и мелководный. Но если у глубоководного вида разнообразие аллелей осталось прежним по сравнению с историческими образцами, то у мелководного помимо своих «родных» аллелей обнаружены почти все аллели глубоководного вида. Мелководный вид постепенно ассимилирует в своем генофонде аллели глубоководного собрата. Так что его вымирание становится неочевидным с философских позиций – вид исчезнет, но гены его останутся.
Все эти озера образовались при таянии ледников около 10 тыс. лет назад. Вскоре эволюция развела специализированные формы по разным экологическим нишам. На этом этапе симпатрического видообразования сформировалась презиготическая изоляция: формы не скрещивались в силу различий в сроках и местах нереста. Этого оказалось достаточно, чтобы появились морфологически различимые виды. Но, как только неблагоприятная обстановка подорвала ресурсы одной из ниш – глубоководной, виды начали обратное схождение. Десяток тысяч лет естественной эволюции перечеркнут полувековым антропогенным вмешательством. Соотношение скоростей естественной и антропогенной эволюции поражает и настраивает на невеселые размышления. Как быстро можно разрушить долгую и кропотливую работу природы!
Но эмоции не должны умалить двух замечательных выводов швейцарско‑канадского исследовательского коллектива: эволюция до какой‑то степени обратима, и вымирание видов не всегда означает их совершенное исчезновение. Эволюция оказалась податливым процессом, а вид, точнее его генетические рудименты, может продолжить существование в генофонде близкого вида. И опять мы возвращаемся к началу этой главы – к рассуждению о сущности биологического вида, о зыбкости и неопределенности формального определения, хотя именно вид, дискретный и стабильный, составляет основу биологического разнообразия природы.
Глава 7
Переходные формы
Проблема переходных форм беспокоила Дарвина, настораживала его сторонников и неизменно радовала оппонентов. Забегая вперед, скажем, что сейчас для экспертов‑биологов эта проблема видится совсем с другой стороны, хотя все еще настораживает малоинформированных приверженцев эволюции и, как и прежде, возбуждает ее противников. В этой главе мы разберем, почему изменилось отношение биологов к переходным формам, а эмоции неспециалистов оставим за скобками.
Переходные формы – это те, что выстраиваются в ряд постепенных переходов от предков к потомкам. Если известно, какой облик имел отдаленный предок, то переходными будут любые наборы промежуточных признаков между ним и потомками. На основе филогенетического дерева можно предсказать, какие промежуточные формы могли существовать (и поэтому могут быть найдены), а какие нет. В соответствии с научным методом сбывшиеся предсказания подтверждают теорию. Например, зная строение динозавров и птиц, можно предсказать кое‑какие морфологические черты переходных форм между ними. Мы прогнозируем возможность найти останки животных, подобных рептилиям, но с перьями, или останки животных, подобных птицам, но с зубами и длинным хвостом. При этом мы предсказываем, что не будут найдены переходные формы между птицами и млекопитающими, например ископаемые млекопитающие с перьями или птицеподобные ископаемые с костями среднего уха как у млекопитающих. Иными словами, эволюционное дерево покажет нам, где и что искать, а чего не может быть и чего искать не стоит.
Кроме того, к переходным относят предковые формы со смешанными признаками дочерних групп. Если дочерние группы происходят напрямую от какого‑то общего предка, то в его облике соберутся некоторые (не обязательно все) примитивные признаки. Другие его признаки могут оказаться, напротив, такими же продвинутыми, как у того или иного потомка. Такую форму можно тоже называть переходной – она совмещает в себе признаки нескольких линий в их примитивном состоянии. Впрочем, подобная ископаемая форма, скорее всего, при ближайшем рассмотрении окажется не прямым предком дочерних групп, а одним из рано ответвившихся и мало измененных потомков этого предка (потому что вероятность найти чьего‑либо прямого предка в очень неполной палеонтологической летописи невелика). Такая форма может служить хорошим портретным приближением к общему предку.
Переходными называют также и виды с промежуточным состоянием сложного признака (если в качестве конечного состояния рассматривать то, что наблюдается у современных животных; с точки зрения наших потомков, нынешняя форма будет промежуточной по сравнению с их обновленным, изменившимся миром).
Суть проблемы переходных форм Дарвин обрисовал так. Если эволюция идет постепенно посредством отбора все более совершенных форм, то мы, казалось бы, должны повсюду видеть ряды бесконечных плавных переходов между формами. В действительности же мы чаще видим дискретные виды, которые либо совсем не могут скрещиваться между собой, либо делают это с трудом и неохотно. В крайнем случае, если ныне живущие «усовершенствованные» виды, победив в конкуренции, вытеснили своих менее приспособленных предков, эти последние должны бы найтись в ископаемой летописи. Пусть современные виды оказались удачливее и выносливее и пусть они теперь расселились повсюду, но ведь когда‑то и их предки были победителями и проживали вольготными царями на своей территории. Это значит, что их останки должны во множестве захорониться, обратиться со временем в фоссилии (окаменеть) и стать достоянием палеонтологов. Но у Дарвина в середине XIX века было мало примеров ископаемых переходных форм.
Другая часть проблемы – это постепенное образование сложного признака. Казалось бы, глаз видит только потому, что в нем все части превосходно подогнаны друг к другу (на самом деле не все и не превосходно, но это уже мелочи). Легкие вдыхают и выдыхают, потому что вся грудная клетка сконструирована как совершенный вакуумный насос. Конструкция уха легка, изящна и математически технична, и потому ухо служит нам, позволяя улавливать мельчайшие оттенки голосовых эмоций, ориентироваться в пространстве и строить звуковые гармонии. Птичье перо – легкая и прочная маховая поверхность; без набора таких совершенных, упорядоченно налегающих друг на друга поверхностей птица не полетит…
Действительно, обретение четко видящего глаза – дело, кажется, исключительно трудное. Ведь его нужно было собирать постепенно, имея в самом начале лишь набор клеток, фиксирующих свет, а в итоге получить точный оптический прибор. И на каждом этапе эволюции этот протоглаз должен был с пользой служить животному, чтобы эволюция не бросила дело на полпути.
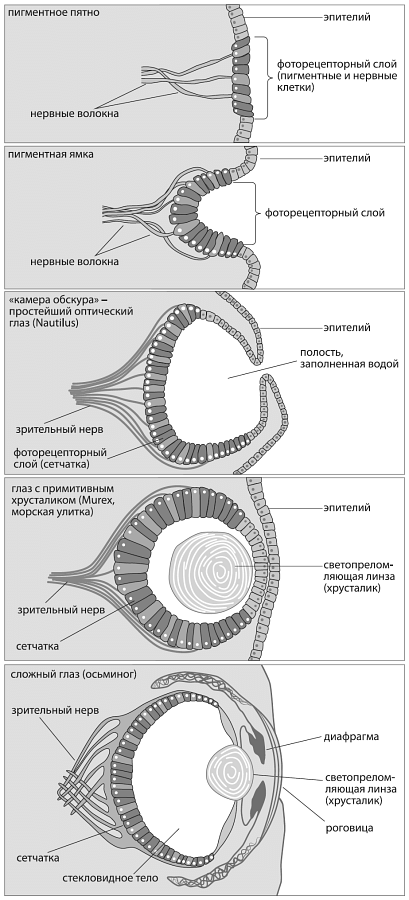
Схема строения глаз современных моллюсков разной степени сложности – от простейшего светочувствительного пятна (у некоторых брюхоногих; вверху ) до глаза, сравнимого по сложности с человеческим (у осьминога; внизу ) – наглядно показывает несостоятельность утверждений, будто такие сложные структуры, как глаза, не могли возникнуть постепенно. Каждое небольшое усовершенствование в этом ряду дает более четкую картинку, а значит, могло быть поддержано отбором. Из Science, Evolution, and Creationism // Washington, D. C.: The National Academies Press.
Представляя мысленно этот путь, неволей задумаешься, зачем бы понадобились животным все эти органы в полусобранном виде. Зачем нужен полуглаз, регистрирующий лишь нечеткие тени, негодное для полета перо, плохо слышащее ухо, дыхательные пузыри, не способные как следует вдохнуть? Стоит ли эволюции тратить на эти приспособления силы и время, если конечный результат неизвестен? А если и вправду был пройден весь путь от пигментного светочувствительного пятна до совершенного глаза, то должны быть переходные формы с полузрячими недоглазами, нелетающими перьями, плохо работающими легкими и тому подобными несовершенными органами. Существовали ли такие формы?
В «Происхождении видов», где этот вопрос обозначен, на него дан четкий теоретический ответ – да, должны были существовать и существовали! – и приведены примеры. Теперь нам известно гораздо, гораздо больше превосходных примеров – переходных форм с такими как будто «недоделанными» морфологическими конструкциями, которые тем не менее прилежно служили своим обладателям.
–––––
Немного о глазах
В действительности глаз не так уж трудно сделать, если в хозяйстве имеются светочувствительные белки (а они есть уже у одноклеточных) и производящие эти белки фоторецепторные клетки (которые появились уже у древнейших животных). Эволюция справлялась с этой задачей неоднократно, каждый раз изобретая особый орган и конструируя его постепенно из подручных материалов (тканей и клеток) согласно нуждам разных животных, но никогда не заглядывая вперед и «заботясь» исключительно о мелких сиюминутных усовершенствованиях.
Глазки двустворчатого моллюска гребешка Свифта, кубомедузы, рыбы‑брызгуна, трилобита – все это совсем разные органы зрения, помогающие животным улавливать свет и различать объекты в характерных для животного условиях. Это примеры параллельного появления сложного органа – глаза.
Глазастое существо на цветной вклейке (см. иллюстрации в конце книги) – гребешок Свифта (Swiftopecten swifti), двустворчатый моллюск размером 10–12 см. По краю мантии у него растут щупальца – органы осязания – и многочисленные мелкие глазки – органы зрения. Глаза гребешка, разумеется, совсем другие, чем у нас, и не связаны с мозгом (его просто нет, есть нервные ганглии), но все же они не так‑то просты. Они представляют собой пузырьки из прозрачного эпителия, заднюю сторону которого выстилает слой светочувствительных клеток, а за ними пигментный слой и так называемое зеркальце. В пузырьке есть светопреломляющая линза – хрусталик. За счет отражения света от зеркальца глазки переливаются чудесным зеленым цветом. С помощью своих глазков гребешки могут видеть лишь на небольшом расстоянии. Только когда злейший враг гребешков – морская звезда – приблизится к нему почти вплотную, он, хлопая створками, совершает прыжок – обращается в бегство. Эти маленькие глазки всегда настороже и предупреждают своего хозяина об опасности.

Кубомедуза.
У медуз (представителей типа кишечнополостных, довольно просто устроенных животных) тоже могут развиваться глаза. Так, кубомедузы – группа с более или менее квадратным куполом – в ходе эволюции обрели зрение. Глаза кубомедуз устроены очень необычно. Они сидят на специальных выростах – ропалиях, их у кубомедуз всего четыре, по числу сторон купола. На каждом ропалии по шесть глаз – четыре простых и два посложнее. Они сконструированы по схеме любого сложного глаза: светочувствительный слой, пигментный слой, роговица и хрусталик. На одном из шести глаз ропалия есть и диафрагма. Изменение освещенности приводит к ее сокращению или расширению: так глаз отвечает на световой сигнал. Эти глаза сконструированы таким образом, чтобы видеть только крупные объекты. Всякую ненужную мелочь медуза просто не замечает. У кубомедуз сигнал от глаз по чувствительным нейронам поступает прямиком к моторным нейронам купола и щупалец. Поэтому, чуть завидев крупный объект, медуза сразу же реагирует: ускоряется и поворачивает. Кубомедузе не нужен анализирующий посредник – мозг или хотя бы нервный ганглий: она не тратит время на раздумья – она видит и действует (Skogh et al., 2006).
Глаза трилобитов – вымерших членистоногих, во множестве населявших моря 530–252 млн лет назад – были и вовсе уникальными. Глаз трилобита фасеточный, как у насекомых: он собран из множества отдельных линз. Линзы, как и у других глазастых членистоногих, прозрачные. Но в отличие от всех без исключения животных они сделаны не из белков, а построены из сверхпрозрачного минерала кальцита. Такой глаз подобен стеклянным очкам, навсегда приросшим к глазу. Так что трилобит обладал в буквальном смысле каменным взором. Каждая линза была строго ориентирована по основной оси светопреломления. Двояковыпуклые линзы фокусировали свет на сетчатке (недавно у трилобитов обнаружены следы пигментных клеток под кальцитовыми линзами), от которой отходили нервы. Линзы, однако, были сильно выпуклыми, почти шарообразными, так что четкость изображения страдала от сферической аберрации. Но трилобиты справились с этой проблемой. Свет фокусировался в точке с помощью тонкой фигурной вставки высокомагнезиального кальцита, имеющего другой коэффициент преломления. Такие линзы с такой же точно фигурной формой, как и в глазах трилобитов, применяются в современных телескопах. Можно утверждать, что естественный отбор решил задачу конструирования современных оптических устройств гораздо раньше Декарта и Гюйгенса, устранивших сферическую аберрацию в телескопах. Эволюция глаз у трилобитов хорошо документирована: от простых множественных двояковыпуклых линз к немногочисленным и почти шарообразным с уплотненной корректирующей вставкой. И те и другие глаза помогали животному определить в воде расстояние до объекта и рассмотреть его издалека. Разница, по‑видимому, состояла в степени детальности изображения и дальнозоркости зрения.
–––––
Таким образом, проблема переходных форм распадается на три вопроса. Первый из них – почему в современном мире не так уж много постепенных переходов между видами, почему большинство видов четко отделяются один от другого. Этот вопрос мы уже обсуждали в главе 6. Второй вопрос можно сформулировать так: почему в ископаемой летописи мало вымерших переходных форм, некогда вытесненных более приспособленными конкурентами? И наконец, последний: как может постепенно сформироваться сложный орган, который кажется полезным только в своей законченной форме? Два последних вопроса мы рассмотрим ниже.








