 2020-06-12
2020-06-12 197
197
Для понимания того, куда идет эволюция человека в наши дни (и идет ли она вообще куда‑то), важно иметь в виду, что практически все наши поведенческие и психологические особенности сильно зависят от генов, а значит, могут эволюционировать под действием отбора здесь и сейчас. Как ни удивительно, сказанное справедливо не только для особенностей темперамента и для предрасположенности к психическим заболеваниям, но и для таких «высокоуровневых» признаков, как число лет, потраченных человеком на учебу. Анализ ДНК сотен тысяч людей европейского происхождения, проведенный в 2016 году, позволил выявить 74 участка генома, вариации в которых достоверно коррелируют с уровнем полученного человеком образования. Значительная часть идентифицированных генов работает в мозге, в том числе во время внутриутробного развития, а мутации многих из них влияют на когнитивные способности. Называть их «генами образования», возможно, не вполне корректно, поскольку их связь с уровнем образования слабая и непрямая – однако она реально существует. Исследование подтвердило, что применение современных методов сравнительной геномики к очень большим выборкам людей позволяет выявить генетический базис даже таких признаков, которые обычно считаются не врожденными.


На первый взгляд может показаться, что уровень образования (традиционно измеряемый числом лет, потраченных человеком на учебу) – типичный пример ненаследственного признака. На него, казалось бы, должны влиять только «факторы среды» в широком смысле: материальная обеспеченность семьи, развитость системы образования в стране, доступность образования для разных социальных групп и т. д. Но это впечатление обманчиво. Если немного подумать, становится ясно, что на уровень образования вполне могут влиять также и признаки, сильно зависящие от генов, – такие, например, как когнитивные способности, целеустремленность или открытость новому опыту.
Еще в 1980‑е годы анализ больших выборок близнецов и их родителей показал, что уровень образования имеет высокую наследуемость (напомним, что наследуемость, которую не следует путать с наследственностью, – это доля изменчивости признака, объясняемая генами, а не средой; подробнее о наследуемости психологических и поведенческих признаков рассказано в нашей книге «Эволюция человека»). Изменчивость по уровню образования как минимум на 20 % определяется генетическими различиями между людьми, а для некоторых выборок были получены значения наследуемости порядка 70 % (в среднем по всем выборкам – примерно 30–40 %; см. рис. 39.1).
Заметим, к слову, что высокие показатели наследуемости таких признаков, как уровень образования или IQ, чаще наблюдаются в выборках людей с высоким социально‑экономическим статусом. У людей с низким статусом наследуемость подобных признаков нередко оказывается ниже (Turkheimer et al., 2003; Hart et al. 2013). Этот пример «взаимодействия генов и среды», скорее всего, связан с тем, что высокий социально‑экономический статус позволяет минимизировать роль случайных факторов в жизни человека. Та фенотипическая изменчивость, которая остается в выборке после устранения роли случайности, при ближайшем рассмотрении оказывается связана в основном с генетическими различиями между людьми. Мы уже использовали подобную логику выше, когда пытались объяснить, почему у видов, заботящихся о потомстве, отбор, возможно, работает эффективнее, чем у видов, бросающих многочисленное потомство на произвол судьбы (см. Исследование № 19). Как забота о потомстве, так и высокий социально‑экономический статус сводит к минимуму элемент случайности в наблюдаемой изменчивости по тем или иным признакам (будь то выживаемость, репродуктивный успех или уровень образования), тем самым выставляя на первый план генетические различия между индивидами.
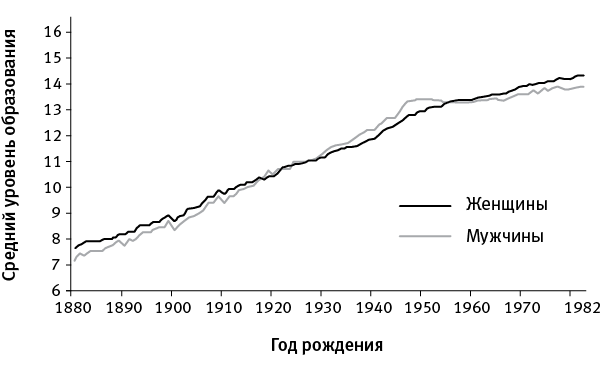
рис. 39.1. Средний уровень образования жителей США (измеряемый по числу лет, затраченных на обучение) в зависимости от года рождения. Видно, что данный показатель неуклонно рос все последнее столетие. Этот рост, очевидно, связан с социальными, культурными и экономическими изменениями, а вовсе не с генетикой. Тем не менее внутри каждой когорты (совокупности людей одного возраста) существует вариабельность по уровню образования, которая на 30–40 % определяется генами. По рисунку с сайта whitehouse.gov.
Но вернемся к образованию. Приведенные значения наследуемости (от 20 до 70 %) отражают общий масштаб генетического вклада в изменчивость по признаку «уровень образования». Гораздо труднее найти конкретные гены, влияющие на этот признак. Дело в том, что число лет обучения – признак «высокоуровневый» в том смысле, что гены влияют на него лишь опосредованно, через множество промежуточных этапов (в отличие, например, от способности воспринимать определенный запах или отличать зеленый цвет от красного – здесь путь от гена к признаку простой и короткий).
На подобные высокоуровневые поведенческие и психологические признаки могут влиять одновременно сотни, если не тысячи разных генов (полиморфных локусов), причем вклад каждого отдельного гена может быть исчезающе мал. В таком случае близнецовый анализ и сравнение родителей с детьми покажут высокую наследуемость признака, но все попытки обнаружить связь между признаком и конкретными аллелями окажутся безуспешными. То есть мы будем знать, что признак сильно зависит от генов, но не сумеем выяснить, от каких именно.
Чтобы преодолеть это затруднение, необходим анализ громадных выборок из десятков и сотен тысяч людей. Чем больше выборка, тем более слабые генетические влияния могут быть с ее помощью обнаружены. Такие исследования стали возможны лишь в последние годы – благодаря прочтению генома человека, развитию и удешевлению молекулярных технологий, а также накоплению генотипических и фенотипических данных по разным человеческим популяциям, собранных по стандартизированным методикам.
В 2013 году были опубликованы результаты первой успешной попытки найти в человеческом геноме конкретные гены, связанные с уровнем образования. Выборка из 126 559 индивидов позволила выявить три гена, каждый из которых достоверно, хотя и очень слабо, ассоциирован с продолжительностью обучения. Объем усилий, понадобившихся для получения этого результата, можно примерно оценить по числу авторов статьи – их около двухсот человек (Rietveld et al., 2013).
В 2016 году в журнале Nature вышла новая статья, в которой еще более внушительный коллектив ученых со всего мира (один только список институтов и университетов, чьи сотрудники приняли участие в исследовании, насчитывает 186 пунктов) сообщил о результатах изучения выборки из 293 723 человек европейского происхождения (Okbay et al., 2016). Фактически был проведен метаанализ, то есть обобщены данные, полученные по единой методике множеством научных коллективов в разных странах. Средняя продолжительность обучения вошедших в выборку людей составляет 14,3 года.
У всех людей, данные по которым были использованы, уровень образования регистрировался в возрасте не менее 30 лет, а генотипирование проводилось по 9,3 млн однонуклеотидных полиморфизмов («снипов»), в совокупности отражающих значительную часть всей генетической вариабельности человечества. Поскольку подавляющее большинство из 3 млрд нуклеотидов в человеческом геноме консервативны, то есть одинаковы у всех (или почти всех) людей, нет необходимости анализировать каждый нуклеотид – достаточно ограничиться только самыми вариабельными.
Исследование представляло собой так называемый полногеномный поиск ассоциаций (GWAS, Genome‑Wide Association Study). У этой методики много тонкостей и подводных камней. Одна из главных проблем связана с популяционной стратификацией. Это значит, что исследуемая популяция, возможно, подразделена на части (субпопуляции), различающиеся по частотам встречаемости каких‑то генов и признаков. Из‑за этого возникает опасность обнаружить ложные ассоциации. Иными словами, мы можем подумать, что данный аллель тесно связан с данным признаком, хотя на самом деле просто случайно так совпало, что в одной из субпопуляций и данный аллель, и данный признак встречаются с повышенной частотой. Это называют «проблемой гена китайских палочек». Название происходит от следующей притчи. Якобы один генетик решил выяснить, какие гены влияют на склонность есть палочками. Он попросил своих студентов (среди которых были индивиды европейского и азиатского происхождения) сообщить ему, как часто они пользуются палочками во время еды. Затем он генотипировал ответивших и провел поиск ассоциаций. Обнаружился локус, строго коррелирующий с использованием палочек. Генетик тут же опубликовал статью, в которой сообщил об открытии гена SUSHI (Successful Use of Selected Hand Instruments). Спустя пару лет выяснилось, что SUSHI на самом деле представляет собой один из генов комплекса гистосовместимости (они очень вариабельны и играют важную роль в работе иммунной системы) и один из аллелей этого гена намного чаще встречается у азиатов, чем у европейцев. Разумеется, этот ген не имеет ни малейшего отношения к использованию палочек. Однако, поскольку в азиатской культуре данное действие распространено шире, чем в европейской, GWAS выявил сильную и достоверную ассоциацию, не имеющую биологического смысла (Hamer, Sirota, 2000).
Чтобы справиться с проблемой «гена китайских палочек», разработаны две группы методов. Первая основана на анализе данных по отдельным семьям: сравнивают родителей и детей или родных братьев и сестер, чтобы проверить, сохраняется ли ассоциация внутри отдельно взятой семьи. «Гены китайских палочек» не выдерживают такой проверки. Вторая группа методов связана с анализом неравновесного сцепления генов: если интересующий нас аллель чаще, чем ожидается при независимом наследовании, встречается в комбинации с определенными аллелями других, произвольно выбранных генов, то есть основания заподозрить, что мы имеем дело с «геном китайских палочек».
Ученые использовали эти и целый ряд других методов, чтобы очистить полученные результаты от ложноположительных сигналов. В итоге удалось выявить 74 полиморфных локуса, каждый из которых с большой вероятностью действительно влияет, тем или иным образом, на продолжительность обучения. Три локуса, найденные в предыдущем исследовании, были повторно выявлены в новом.
Как и следовало ожидать, все эти гены влияют на уровень образования слабо. Эффект каждого по отдельности соответствует 3–9 неделям учебы и объясняет от 0,01 до 0,035 % изменчивости по признаку «продолжительность обучения». Причем суммарный эффект всех 74 генов меньше суммы их индивидуальных эффектов.
В распоряжении исследователей имелись сведения не только о продолжительности обучения протестированных индивидов, но и о других фенотипических признаках, в том числе связанных с работой мозга. Это позволило оценить генетические корреляции, то есть то, в какой мере гены, влияющие на уровень образования, влияют также и на другие признаки.
Получилось, что генетические варианты, способствующие долгой учебе, коррелируют также с хорошими умственными способностями, большим объемом мозга и почему‑то с повышенным риском биполярного расстройства. Обнаружены также крайне слабые, но все же достоверные положительные корреляции с риском шизофрении и ростом. Для этих же полиморфизмов выявлены и достоверные отрицательные корреляции: с невротизмом, риском болезни Альцгеймера, а также с массой тела.
Функциональный спектр этих 74 генов оказался весьма показательным. Среди них резко повышена доля генов, участвующих в развитии и работе мозга. Мутации в них часто ведут к умственной отсталости, уменьшению объема мозга, повышают риск болезни Альцгеймера и других нарушений когнитивных функций. Одни экспрессируются в развивающемся мозге у эмбрионов, регулируя деление клеток‑предшественников нейронов, миграцию молодых нейронов и рост неокортекса, другие играют роль в синаптической пластичности (в том числе – в образовании дендритных шипиков и новых синапсов) в течение всей жизни. Синаптическая пластичность, как известно, лежит в основе долговременной памяти, без которой невозможно чему‑то научиться.
Тесная связь «генов образования» с работой мозга представляется вполне логичной. Вряд ли кого‑то удивит, что индивиды с хорошо развитым мозгом, при прочих равных условиях, проявляют больше склонности к учебе. И это, кстати, говорит о том, что влияние данных генов на учебу не такое уж и косвенное. Ведь могло оказаться и так, что какой‑нибудь ген повышает уровень образования, поскольку влияет на свойства таза и позвоночника, помогающие спокойно сидеть за партой. Но нет: «гены образования» влияют на мозг, а не на то, на чем мы сидим.
Исследователи, впрочем, считают, что называть выявленные ими гены «генами образования» некорректно. Во‑первых, уровень образования определяется средовыми факторами в несколько большей степени, чем генетическими. Но это справедливо для большинства психологических и поведенческих признаков человека. Во‑вторых, влияние отдельных генов на рассматриваемый признак невелико. Это, правда, тоже общее место в генетике поведения. В‑третьих, аллельные варианты генов могут влиять на признак «продолжительность обучения» не непосредственно, а через ряд промежуточных фенотипов. Конечно, это тоже справедливо для большинства связей между генами и поведенческими признаками у приматов. Чтобы проиллюстрировать последнее соображение, исследователи рассчитали, что выявленная связь между генами и продолжительностью учебы на 23–42 % объясняется влиянием этих генов на умственные способности, а еще 7 % корреляции можно списать на влияние этих же генов на личностную характеристику «открытость новому опыту». Насколько приведенные аргументы нетривиальны и насколько из них действительно следует, что не надо называть эти гены «генами образования», судить читателю. На наш взгляд, это просто дань политкорректности.
Авторы также отмечают, что из их результатов вовсе не следует, будто влияние 74 генов на длительность учебы является некой раз и навсегда заданной, неизменной величиной. Напротив, имеющиеся данные позволяют утверждать, что степень влияния этих и других генов на признак «длительность учебы» зависит от средовых условий. Она различна для разных стран и меняется со временем в одной и той же стране. Тут нет ничего удивительного: наследуемость признаков – величина непостоянная. Она может и должна меняться в зависимости от социально‑экономических и других условий (мы подробно рассказали об этом в книге «Эволюция человека»).
Так или иначе, исследование показало, что если очень постараться и набрать побольше данных, то можно найти конкретные гены, влияющие на полигенный признак, даже в том случае, если вклад каждого отдельного гена ничтожно мал. Кроме того, работа в очередной раз продемонстрировала, что даже сложные поведенческие признаки, которые традиционно считаются ненаследственными (приобретенными), в действительности могут иметь значительную наследственную составляющую.
Исследование № 40
«Гены образования» отсеиваются отбором
Итак, мы знаем, что в современных человеческих популяциях уровень полученного образования сильно зависит от генов: выявлено 74 полиморфных гена, влияющих на этот признак. Причем, как выясняется, уровень образования отрицательно коррелирует с дарвиновской приспособленностью: образованные люди хуже размножаются. Это указывает на возможный отбор против «генов образования». Новое исследование, основанное на данных по 110 000 исландцев, показало, что «гены образования» действительно подвергаются отрицательному отбору. Эти аллели, многие из которых коррелируют также с высоким интеллектом, крепким здоровьем и долгой жизнью, снижают приспособленность (число оставляемых потомков) независимо от того, реализовал ли человек обусловленную ими склонность к получению хорошего образования. Исследование подтвердило опасения, что эволюция современного человечества направлена в сторону ухудшения генетического базиса признаков, связанных с интеллектом. Социально‑культурное развитие пока с лихвой компенсирует генетическую деградацию, но со временем ее последствия могут стать существенными.


Генетическая деградация человечества и ее возможные механизмы. Идею о том, что в современном цивилизованном обществе (в отличие от общества «варварского») эволюция направлена совсем не в ту сторону, в какую нам хотелось бы, впервые подробно изложил и обосновал величайший генетик‑эволюционист XX века Рональд Фишер в заключительных главах книги «Генетическая теория естественного отбора» (1930 г.).
Исследования в этой области продвигались медленно, в том числе по причинам, не имеющим отношения к науке. Тем не менее к настоящему времени накопилось немало данных, косвенно подтверждающих, что генетический базис признаков, связанных с физическим и умственным здоровьем, размывается и деградирует, особенно у жителей развитых стран. Генетики обсуждают два основных механизма такой деградации (Woodley of Menie, 2015).
Первый механизм – ослабление очищающего отбора против мутаций, нарушающих работу тех или иных органов и систем организма, в том числе иммунной системы и мозга. Раньше такие мутации снижали приспособленность (их носители оставляли в среднем меньше здорового потомства) и потому вычищались отбором. Однако развитие медицины, социального обеспечения и других благ цивилизации сделало эти мутации менее вредными в эволюционном смысле, а то и вовсе безвредными (нейтральными). Поэтому они стали свободно накапливаться в генофонде. Развитие и работа коры головного мозга зависят от слаженной работы огромного количества генов, что делает наши когнитивные способности особенно уязвимыми перед мутационной деградацией (Crabtree, 2013).
Второй механизм – отрицательный отбор по признакам, помогающим добиваться успеха в жизни. Раньше такие признаки повышали дарвиновскую приспособленность, а в современном обществе они ее скорее снижают – например, в силу того, что талантливые, образованные и экономически успешные люди стали откладывать рождение детей на потом. Так, есть данные, согласно которым в США и других богатых странах люди с высокими показателями интеллекта оставляют меньше потомства (Lynn, 1999). Есть также основания полагать, что в развитых странах после промышленной революции направленность отбора по признакам, связанным с социальным и экономическим успехом, стала меняться на противоположную. Скажем, если раньше умелый сапожник и сметливый торговец оставляли больше потомства, чем их криворукие и бестолковые конкуренты, то начиная примерно с середины XIX века все стало наоборот (Skirbekk, 2008). Возможные причины – развитие медицины и социального обеспечения (дети бедных и неудачливых родителей перестали массово погибать в младенчестве), а также средств планирования семьи (индивиды, ориентированные на жизненный успех, получили возможность откладывать рождение детей).
Впрочем, до сих пор в распоряжении ученых были в основном лишь косвенные свидетельства генетической деградации человечества. Не хватало прямых доказательств того, что гены, способствующие развитию ценимых нами психологических и когнитивных признаков, действительно подвержены отрицательному отбору и что этот отбор эффективен (то есть ведет к снижению частоты встречаемости соответствующих аллелей). И вот, благодаря развитию методов сравнительной геномики и созданию больших баз данных по медицинской генетике, такие доказательства наконец появились, с чем нас всех можно поздравить.
Комплексный показатель генетической предрасположенности к получению образования. Исландские генетики изучили связь между генами, влияющими на уровень образования, и дарвиновской приспособленностью на примере выборки, включающей значительную часть (примерно половину) всех исландцев, родившихся с 1910 по 1975 год (Kong et al., 2017).
Уровень полученного образования, измеряемый числом лет, потраченных на учебу, – удобный для изучения фенотипический признак, тесно связанный с когнитивными способностями и такими особенностями психики, которые мы обычно считаем достоинствами (целеустремленность, тяга к знаниям, способность планировать свою жизнь, открытость новому опыту). Установлено, что уровень образования в современных человеческих популяциях имеет высокую наследуемость. Различия между людьми по этому признаку в среднем на 30–40 % объясняются генами. Остальные 60–70 % – результат различающихся условий среды и онтогенетического шума (см. Исследование № 39).
В 2016 году на выборке из 20 000 американцев впервые удалось показать, что генетический базис образования подвергается отрицательному отбору (Beauchamp, 2016). Работа исландских генетиков, опубликованная в начале 2017 года, не только подтвердила этот вывод на более представительной выборке, но и продемонстрировала реальное снижение частоты встречаемости аллелей, способствующих получению образования, в течение XX века.
Обсуждаемая работа во многом основывается на исследовании, описанном выше (см. Исследование № 39), в котором анализировалась выборка, включающая 293 724 лица европейского происхождения. Анализ этой выборки позволил выявить 74 локуса (участка генома), достоверно влияющих на уровень образования. Исландские генетики использовали полученные ранее данные, чтобы вывести формулу для вычисления «комплексного показателя генетической предрасположенности к получению образования» – POLYEDU (educational attainment polygenic score). Этот показатель с точностью, зависящей от размера изучаемой выборки, отражает совместный вклад множества генов (в идеале – всех в геноме) в предрасположенность данного человека к получению образования. Показатель POLYEDUскладывается из индивидуальных весов, которые были рассчитаны для каждого из 620 000 генетических маркеров (полиморфных локусов) в зависимости от того, как аллельные варианты этих локусов соотносятся с уровнем образования. Если наличие аллеля ассоциировано с немного повышенным уровнем образования, то такому аллелю придается небольшой положительный вес, и т. д. Ученые перепробовали несколько способов вычисления POLYEDU и убедились, что методические тонкости мало влияют на итоговые результаты.
Чтобы избежать лишних трудностей с интерпретацией результатов, при выведении формулы для POLYEDU из исходной выборки в 293 724 индивида были исключены все исландцы, которые туда входили (49 970), а вместо них добавлены 111 349 человек из британской базы данных UK Biobank. Увеличение выборки привело к тому, что индивидуальных локусов, достоверно коррелирующих с уровнем образования, стало уже не 74, а 120.
POLYEDU, вычисляемый по полученной формуле, позволяет объяснить 3,74 % изменчивости по уровню образования. Иными словами, этот показатель характеризует примерно одну десятую часть всего генетического базиса данного признака (поскольку весь генетический базис объясняет 30–40 % изменчивости). Если же формулу для POLYEDU вывести на основе меньшей выборки – без учета данных из UK Biobank, – то получившийся показатель будет объяснять лишь 2,52 % изменчивости. По‑видимому, для того чтобы рассчитать вес каждого аллеля еще точнее – так, чтобы итоговый показатель объяснял бóльшую долю генетически обусловленной вариабельности по уровню образования, – нужны данные не по сотням тысяч, а по миллионам людей. Показатель POLYEDU нормировали таким образом, чтобы его среднее значение равнялось нулю, а стандартное отклонение – единице, и затем измеряли его в условных единицах.
«Гены образования» снижают приспособленность. По полученной формуле ученые вычислили POLYEDU для 109 120 исландцев, родившихся c 1910 по 1975 год. Индивидуальные значения POLYEDU были сопоставлены с тремя репродуктивными показателями:
1) Общее количество потомков, оставленных человеком за всю жизнь, не считая умерших в младенчестве. Этот показатель близок по смыслу к дарвиновской приспособленности. Здесь нужно пояснить, что в современных развитых обществах смертность и в детском, и в репродуктивном возрасте настолько низка, а системы социального обеспечения настолько хорошо развиты, что приспособленность зависит лишь от количества детей, но не от их «качества». Практически все дети доживают до зрелости и получают возможность поучаствовать в размножении – независимо от того, крепкое ли у них здоровье и хорошо ли о них заботились родители.
2) Возраст родителя в момент рождения первого ребенка.
3) Средний возраст родителя при рождении потомства.
Результаты анализа приведены в таблице:
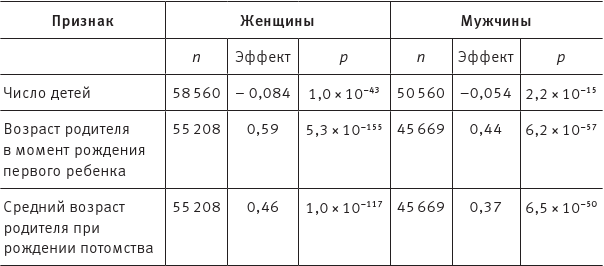
Таблица показывает корреляцию между POLYEDU и тремя репродуктивными показателями. Числа в столбце «Эффект» означают, что увеличение POLYEDU на единицу сопровождается изменением данного признака на указанную величину. Возраст измеряется в годах, n – число людей в выборке, p – мера достоверности обнаруженной взаимосвязи (проще говоря, это вероятность того, что эффект такой силы обнаружился бы в выборке чисто случайно, если бы никакой связи на самом деле не было). Из таблицы следует, что у женщин увеличение POLYEDU на единицу соответствует уменьшению количества детей на 0,084 и увеличению возраста рождения первого ребенка на 0,59 года, а среднего возраста деторождения – на 0,46. Соответствующие значения для мужчин: 0,054, 0,44 и 0,37.
Возможно, результаты покажутся более наглядными, если мы посмотрим на различия между людьми с самой слабой и самой сильной врожденной склонностью к получению образования. Исследование показало, что 2 % женщин с самым низким POLYEDU начинают рожать почти на 3 года раньше и в итоге оставляют на 0,5 ребенка больше, чем 2 % женщин с самым высоким POLYEDU.
Итак, гены, способствующие образованию, достоверно снижают репродуктивный успех у обоих полов. Кроме того, они содействуют сдвигу начала репродукции на более поздний возраст. Эти эффекты у женщин выражены сильнее, чем у мужчин. Показатель «возраст рождения первого ребенка» увеличивается с ростом POLYEDU быстрее, чем «средний возраст деторождения». Это позволяет предположить (а дополнительные расчеты подтвердили это предположение), что «гены образования» снижают плодовитость в основном за счет того, что рождение первого ребенка сдвигается на более поздний возраст.
Более детальный анализ показал, что «гены образования» резко снижают раннюю плодовитость, тогда как на деторождение в позднем возрасте они влияют положительно. Ранний отрицательный эффект сильно перевешивает поздний положительный, так что итоговое число детей оказывается существенно ниже у людей с высоким POLYEDU. Перелом происходит в возрасте около 30 лет. Вплоть до этого возраста лучше размножаются люди с низким POLYEDU. После 30 лет обладатели высокого POLYEDU немного сокращают разрыв по числу детей, но догнать «соперников» уже не могут.
Данное исследование посвящено совокупному влиянию множества генов, а не роли отдельных аллелей. Индивидуальные вклады большинства генов в вариабельность по уровню образования очень малы, поэтому для их изучения размер выборки должен быть больше. Но некоторые аллели ученые все же отметили. Например, один вариант полиморфного локуса rs 192818565 негативно влияет на уровень образования. При этом он подвергается положительному отбору, то есть повышает дарвиновскую приспособленность. Известно также, что присутствие этого аллеля коррелирует с невротизмом и уменьшенным объемом черепной коробки. Как выяснилось, этот явно «плохой» аллель (кому же хочется быть невротиком с маленьким мозгом?) достоверно коррелирует с бóльшим числом детей и ранним началом деторождения. Как справедливо отмечают авторы, «это поразительный пример генетического варианта, который ассоциирован с фенотипом, обычно рассматриваемым как неблагоприятный, и одновременно – с повышенной приспособленностью в эволюционном смысле».
Отбор против «генов образования» не сводится к отбору против образованных людей. Следующий вопрос, который ученые попытались решить, касается механизма влияния POLYEDU на приспособленность. Поскольку известно, что POLYEDU отражается на образовании, а образование негативно сказывается на деторождении, то логично предположить, что отрицательное влияние POLYEDU на репродукцию целиком и полностью объясняется его положительным влиянием на образование. Чтобы проверить, так ли это, исследователи проанализировали совместное влияние двух факторов – POLYEDU и уровня образования – на репродуктивные показатели. Выяснилось, что уровень образования влияет на репродуктивный успех женщин примерно так же, как и POLYEDU, снижая общее число детей (на 0,045 за год обучения) и сдвигая начало репродукции на более поздний возраст (на 4,2 месяца за год обучения). Однако эффекты этих двух факторов нельзя свести друг к другу: они действуют во многом независимо. Если рассчитать влияние POLYEDU на приспособленность с поправкой на уровень образования, то это влияние лишь слегка уменьшается, оставаясь высоко достоверным. Например, если взять только женщин, получивших десятилетнее образование (это обязательный минимум в Исландии), то мы увидим, что внутри этой выборки женщины с высоким POLYEDU родили меньше детей, чем женщины с низким, – несмотря на то, что уровень образования у всех одинаковый. Таким образом, гены, повышающие склонность к получению образования, снижают приспособленность независимо от того, реализовал человек эту склонность или нет. То же самое получается, если рассчитать влияние уровня образования на приспособленность с поправкой на POLYEDU: образование снижает приспособленность женщин независимо от того, насколько выражена у них генетическая предрасположенность к его получению.
У мужчин негативное влияние POLYEDU на приспособленность тоже не сводится к влиянию POLYEDU на образование. Более того, оказалось, что образование само по себе не снижает, а даже немного увеличивает репродуктивный успех мужчин! Каждый лишний год образования прибавляет исландскому мужчине в среднем по 0,011 ребенка. Таким образом, генетическая склонность к получению образования снижает приспособленность мужчин вопреки тому, что образование само по себе повышает репродуктивный успех.
Из этого напрямую следует, что «гены образования», помимо своего влияния на уровень образования, имеют и другие фенотипические эффекты, негативно сказывающиеся на приспособленности. Что же это за эффекты?
Оказалось, что POLYEDU положительно коррелирует с продолжительностью жизни, а также с целым рядом параметров, связанных с физическим и умственным здоровьем. В частности, люди с высоким POLYEDU имеют пониженный уровень триглицеридов в крови, реже страдают ожирением и меньше курят. Кроме того, POLYEDU положительно коррелирует с показателями интеллекта, такими как способность решать задачки из тестов на IQ.

Отрицательное влияние физического здоровья на репродукцию представить себе трудно. Дело тут скорее в интеллекте. Например, логично предположить, что умные люди планируют свою жизнь и стараются не заводить детей слишком рано. Это самый простой и очевидный механизм, посредством которого интеллект может снижать приспособленность, а значит, содействовать естественному отбору на поглупение. Поскольку интеллект положительно коррелирует с физическим здоровьем, такой отбор может попутно делать нас более слабыми и болезненными.
Генетический базис образования действительно ухудшается. Итак, POLYEDU отрицательно коррелирует с дарвиновской приспособленностью. Это значит, что естественный отбор работает против «генов образования». Следовательно, частота встречаемости этих генов в генофонде популяции должна со временем снижаться. Ученые проверили это, сравнив средние значения POLYEDU у исландцев, рожденных в разные годы. Оказалось, что частота встречаемости «генов образования» действительно снижалась в течение всего исследованного отрезка времени (с 1910 по 1975 год). Таким образом, отрицательный отбор по «генам образования» вполне эффективен: эти гены постепенно выбраковываются из генофонда. Наблюдаемая скорость снижения POLYEDU примерно совпадает с ожидаемой на основе данных по плодовитости, приведенных в таблице.
На пути к «идиократии». Поскольку POLYEDU положительно коррелирует с когнитивными способностями, отрицательный отбор по «генам образования» должен вести к поглупению популяции. Чтобы оценить масштаб бедствия, ученые сделали правдоподобное допущение: они предположили, что те влияющие на образование гены, чьи эффекты не учитываются показателем POLYEDU, влияют на приспособленность и IQ так же, как гены, чьи эффекты в POLYEDUотражены. В таком случае получается, что ухудшение генетического базиса интеллекта под действием отрицательного отбора по «генам образования» должно вести к снижению среднего IQ популяции на 0,3 балла за десятилетие. Если такой отбор будет продолжаться много веков подряд, последствия окажутся весьма ощутимыми. Скажем, за 1000 лет среднее значение IQ снизится на 30 баллов, а это, скорее всего, будет означать крах цивилизации.
Правда, реальный уровень IQ людей в XX веке не снижался, а рос за счет социально‑культурных факторов (эта тенденция известна под названием «эффект Флинна»). Средняя скорость роста с 1932 по 1978 год составила 3 балла за десятилетие, что с лихвой перекрывает предполагаемую генетическую деградацию. Но эффект Флинна не имеет отношения к «генам интеллекта» и к биологической эволюции: наблюдаемый рост идет слишком быстро, чтобы пытаться связать его с эволюционной генетикой. Он отражает социальные и культурные процессы, содействующие более полному раскрытию врожденных способностей к интеллектуальному росту. Сами же эти способности, к сожалению, деградируют. И у нас нет оснований надеяться, что все это касается только исландцев: на американцах были получены такие же результаты, а, судя по многим косвенным признакам, похожие процессы идут и в других странах.
Мы не знаем, как долго будет работать эффект Флинна, но вряд ли стоит на него рассчитывать в долгосрочной перспективе. Есть данные, указывающие на ослабление и даже полное исчезновение эффекта Флинна в некоторых популяциях начиная с 1990‑х годов. Впрочем, мы не знаем и того, как долго будет продолжаться отрицательный отбор по образованию и интеллекту. Ведь в ходе антропогенеза те же самые гены, очевидно, подвергались положительному отбору, так что неизвестно, как будет меняться направленность отбора в будущем.
Так или иначе, исследование показало, что опасения Рональда Фишера не были беспочвенными. Может, как он и предполагал, любая цивилизация на определенном этапе своего развития действительно создает условия для отрицательного отбора по генам, сделавшим нас разумными существами, что в конечном счете приводит к коллапсу этой самой цивилизации и (в лучшем случае) впадению в варварство? Если уж на то пошло, не этим ли объясняются парадокс Ферми и молчание космоса?
Мы не знаем. Но если тенденции, выявленные у исландцев, действуют и в других странах (а это почти наверняка так) и если все это будет продолжаться еще несколько веков, то генетическая деградация станет для человечества серьезной проблемой. Чтобы ей противостоять, необходимо побыстрее развивать науку вообще и эволюционную биологию в частности – пока еще есть кому. Разумеется (и к счастью), сегодня речь уже не может идти о варварских методах искусственного отбора, обсуждавшихся основоположниками евгеники. Но есть смысл думать о разработке высокотехнологичных и гуманных методов коррекции неблагоприятных эволюционных тенденций. В том числе – методов, связанных с отбором гамет или ранних эмбрионов, генной терапией и генной инженерией.
Список литературы
1. Марков А. В. и др. 2015. Материнский эффект маскирует адаптацию к неблагоприятным условиям и затрудняет дивергенцию у Drosophila melanogaster // Журнал общей биологии. Т. 76. С. 429–437.
2. Трут Л. Н. 2007. Обретет ли человек нового друга? // Природа. № 6. С. 11–17.
3. Abbot P. et al. 2011. Inclusive fitness theory and eusociality // Nature. V. 471. P. e1–e4.
4. Alcaide M. et al. 2014. Genomic divergence in a ring species complex // Nature. V. 511. P. 83–85.
5. Alexander R. 1974. The evolution of social behavior // Annu Rev Ecol Syst. V. 5. P. 325–383.
6. Baker C. R. et al. 2013. Following gene duplication, paralog interference constrains transcriptional circuit evolution // Science. V. 342. P. 104–108.
7. Beauchamp J. P. 2016. Genetic evidence for natural selection in humans in the contemporary United States // Proc Natl Acad Sci USA. V. 113. P. 7774–7779.
8. Brawand D. et al. 2014. The genomic substrate for adaptive radiation in African cichlid fish // Nature. V. 513. P. 375–381.
9. Buffington S. A. et al. 2016. Microbial reconstitution reverses maternal diet‑induced social and synaptic deficits in offspring // Cell. V. 165. P. 1762–1775.
10. Camperio‑Ciani A. et al. 2004. Evidence for maternally inherited factors favouring male homosexuality and promoting female fecundity // Proc Biol Sci. V. 271. P. 2217–2221.
11. Carazo P. et al. 2014. Within‑group male relatedness reduces harm to females in Drosophila // Nature. V. 505. P. 672–675.
12. Coleman S. W. et al. 2009. Reproductive isolation, reproductive mode, and sexual selection: empirical tests of the viviparity‑driven conflict hypothesis // Am Nat. V. 173. P. 291–303.
13. Condon M. A. et al. 2014. Lethal interactions between parasites and prey increase niche diversity in a tropical community // Science. V. 343. P. 1240–1244.
14. Corral‑López A. et al. 2017. Female brain size affects the assessment of male attractiveness during mate choice // Sci Adv. V. 3. P. e1601990.
15. Crabtree G. R. 2013. Our fragile intellect. Part II // Trends Genet. V. 29. P. 3–5.
16. Daly M., Wilson V. 1988. Evolutionary social psychology and family homicide // Science. V. 242. P. 519–521.
17. Dannemann M. et al. 2016. Introgression of Neandertal‑ and Denisovan‑like haplotypes contributes to adaptive variation in human Toll‑like receptors // Am J Hum Genet. V. 98. P. 22–33.
18. Deschamps M. et al. 2016. Genomic signatures of selective pressures and introgression from archaic hominins at human innate immunity genes // Am J Hum Genet. V. 98. P. 5–21.
19. Diss G. et al. 2017. Gene duplication can impart fragility, not robustness, in the yeast protein interaction network // Science. V. 355. P. 630–634.
20. Endler J. A. 1980. Natural selection on color patterns in Poecilia reticulata // Evolution. V. 34. P. 76–91.
21. Fuentes I. et al. 2014. Horizontal genome transfer as an asexual path to the formation of new species // Nature. 2014. V. 511. P. 232–235.
22. Gasparini C. et al. 2012. Cross‑generational effects of sexual harassment on female fitness in the guppy // Evolution. V. 66. P. 532–543.
23. Ghalambor C. K. et al. 2015. Non‑adaptive plasticity potentiates rapid adaptive evolution of gene expression in nature // Nature. V. 525. P. 372–375.
24. Gibson A. K. et al. 2017. The two‑fold cost of sex: experimental evidence from a natural system // Evol Lett. 2017. V. 1. P. 6–15.
25. Good B. H. et al. 2017. The dynamics of molecular evolution over 60,000 generations // Nature. V. 551. P. 45–50.
26. Grant P. R., Grant B. R. 2006. Evolution of character displacement in Darwin’s finches // Science. V. 313. P. 224–226.
27. Grant P. R., Grant B. R. 2014. Speciation undone // Nature. V. 507. P. 178–179.
28. Gross J., Bhattacharya D. 2010. Uniting sex and eukaryote origins in an emerging oxygenic world // Biol Direct. V. 5. P. 53.
29. Hamer D., Sirota L. 2000. Beware the chopsticks gene // Mol Psychiatry. V. 5. P. 11–13.
30. Harris K. 2015. Evidence for recent, population‑specific evolution of the human mutation rate // Proc Natl Acad Sci USA. V. 112. P. 3439–3444.
31. Hart S. A. et al. 2013. Expanding the environment: gene × school‑level SES interaction on reading comprehension // J Child Psychol Psychiatry. V. 54. P. 1047–1055.
32. Highton R. 1998. Is Ensatina eschscholtzii a ring‑species? // Herpetologica. V. 54. P. 254–278.
33. Hooper D. M., Price T. D. 2015. Rates of karyotypic evolution in Estrildid finches differ between island and continental clades // Evolution. V. 69. P. 890–903.
34. Hooper D. M., Price T. D. 2017. Chromosomal inversion differences correlate with range overlap in passerine birds // Nat Ecol Evol. V. 1. P. 1526–1534.
35. Hsiao E. Y. et al. 2013. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders // Cell. V. 155. P. 1451–1463.
36. Inoue Y. et al. 2017. Evolution of the wheat blast fungus through functional losses in a host specificity determinant // Science. V. 357. P. 80–83.
37. Johnston S. E. et al. 2013. Life history trade‑offs at a single locus maintain sexually selected genetic variation // Nature. V. 502. P. 93–95.
38. Jónsson H. et al. 2017. Parental influence on human germline de novo mutations in 1,548 trios from Iceland // Nature. V. 549. P. 519–522.
39. Keagy J. et al. 2009. Male satin bowerbird problem‑solving ability predicts mating success // Anim Behav. V. 78. P. 809–817.
40. Kleindorfer S. et al. 2014. Species collapse via hybridization in Darwin’s tree finches // Am Nat. V. 183. P. 325–341.
41. Kong A. et al. 2017. Selection against variants in the genome associated with educational attainment // Proc Natl Acad Sci USA. V. 114. P. e727–e732.
42. Kopelman P. 2000. Obesity as a medical problem // Nature. V. 404. P. 635–643.
43. Krakowiak P. et al. 2012. Maternal metabolic conditions and risk for autism and other neurodevelopmental disorders // Pediatrics. V. 129. P. e1121–e1128.
44. Kulmuni J., Pamilo P. 2014. Introgression in hybrid ants is favored in females but selected against in males // Proc Natl Acad Sci USA. V. 111. P. 12805–12810.
45. Lack D. 1947. Darwin’s finches. Cambridge University Press.
46. Lamichhaney S. et al. 2016. A beak size locus in Darwin’s finches facilitated character displacement during a drought // Science. V. 352. P. 470–474.
47. Lamichhaney S. et al. 2018. Rapid hybrid speciation in Darwin’s finches // Science. V. 359. P. 224–228.
48. Langergraber K. E. et al. 2012. Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution // Proc Natl Acad Sci USA. V. 109. P. 15716–15721.
49. Lee S.‑G. et al. 2017. Age‑associated molecular changes are deleterious and may modulate life span through diet // Sci Adv. V. 3. P. e1601833.
50. Levy S. F. et al. 2015. Quantitative evolutionary dynamics using high‑resolution lineage tracking // Nature. V. 519. P. 181–186.
51. Liebers D. et al. 2004. The herring gull complex is not a ring species // Proc Biol Sci. V. 271. P. 893–901.
52. Lukas D., Huchard E. 2014. The evolution of infanticide by males in mammalian societies // Science. V. 346. P. 841–844.
53. Łukasiewicz A. et al. 2017. Kin selection promotes female productivity and cooperation between the sexes // Sci Adv. V. 3. P. e1602262.
54. Lumley A. J. et al. 2015. Sexual selection protects against extinction // Nature. V. 522. P. 470–473.
55. Lynch M. 2010a. Evolution of the mutation rate // Trends Genet. V. 26. P. 345–352.
56. Lynch M. 2010b. Rate, molecular spectrum, and consequences of human mutation // Proc Natl Acad Sci USA. V. 107. P. 961–968.
57. Lynn R. 1999. New evidence for dysgenic fertility for intelligence in the United States // Soc Biol. V. 46. P. 146–153.
58. Makarova K. S. et al. 2005. Ancestral paralogs and pseudoparalogs and their role in the emergence of the eukaryotic cell // Nucleic Acids Res. V. 33. P. 4626–4638.
59. Markov A. V., Kaznacheev I. S. 2016. Evolutionary consequences of polyploidy in prokaryotes and the origin of mitosis and meiosis // Biol Direct. V. 11. P. 28.
60. Martins M. J. F. et al. 2018. High male sexual investment as a driver of extinction in fossil ostracods // Nature. V. 556. P. 366–369.
61. McDonald M. J. et al. 2016. Sex speeds adaptation by altering the dynamics of molecular evolution // Nature. V. 531. P. 233–236.
62. Meirmans S. et al. 2012. The costs of sex: facing real‑world complexities // Q Rev Biol. V. 87. P. 19–40.
63. Nadeau N. J. et al. 2016. The gene cortex controls mimicry and crypsis in butterflies and moths // Nature. V. 534. P. 106–110.
64. Nowak M. A. et al. 2010. The evolution of eusociality // Nature. V. 466. P. 1057–1062.
65. Okbay A. et al. 2016. Genome‑wide association study identifies 74 loci associated with educational attainment // Nature. V. 533. P. 539–542.
66. Ostrovsky A. N. et al. 2015. Matrotrophy and placentation in invertebrates: a new paradigm // Biol Rev Camb Philos Soc. V. 91. P. 673–711.
67. Ozawa H. 2013. The history of sexual dimorphism in Ostracoda (Arthropoda, Crustacea) since the Palaeozoic // Sexual Dimorphism. InTech open.
68. Panchin A. Y. et al. 2014. Midichlorians – the biomeme hypothesis: is there a microbial component to religious rituals? // Biol Direct. V. 9. P. 14.
69. Penz O. K. et al. 2015. Protracted brain development in a rodent model of extreme longevity // Sci Rep. V. 5. P. 11592.
70. Podgornaia A. I., Laub M. T. 2015. Pervasive degeneracy and epistasis in a protein‑protein interface // Science. V. 347. P. 673–677.
71. Poduri A. et al. 2013. Somatic mutation, genomic variation, and neurological disease // Science. V. 341. P. 1237758.
72. Pollux B. J. A. et al. 2014. The evolution of the placenta drives a shift in sexual selection in livebearing fish // Nature. V. 513. P. 233–236.
73. Poutahidis T. et al. 2013. Microbial symbionts accelerate wound healing via the neuropeptide hormone oxytocin // PLoS One. V. 8. P. e78898.
74. Reznick D. N. et al. 2002. Independent origins and rapid evolution of the placenta in the fish genus Poeciliopsis // Science. V. 298. P. 1018–1020.
75. Rietveld C. A. et al. 2013. GWAS of 126,559 individuals identifies genetic variants associated with educational attainment // Science. V. 340. P. 1467–1471.
76. Roberts E. K. et al. 2012. A Bruce effect in wild geladas // Science. V. 335. P. 1222–1225.
77. Romiguier J. et al. 2014. Comparative population genomics in animals uncovers the determinants of genetic diversity // Nature. V. 515. P. 261–263.
78. Saha S. et al. 2009. Advanced paternal age is associated with impaired neurocognitive outcomes during infancy and childhood // PLoS Med. V. 6. P. e40.
79. Sankararaman S. et al. 2014. The genomic landscape of Neanderthal ancestry in present‑day humans // Nature. V. 507. P. 354–357.
80. Scally A., Durbin R. 2012. Revising the human mutation rate: implications for understanding human evolution // Nat Rev Genet. V. 13. P. 745–753.
81. Shendure J., Akey J. M. 2015. The origins, determinants, and consequences of human mutations // Science. V. 349. P. 1478–1483.
82. Shohet A. J., Watt P. J. 2009. Female guppies Poecilia reticulata prefer males that can learn fast // J Fish Biol. V. 75. P. 1323–1330.
83. Simonti C. N. et al. 2016. The phenotypic legacy of admixture between modern humans and Neanderthals // Science. V. 351. P. 737–741.
84. Skirbekk V. 2008. Fertility trends by social status // Demogr Res. V. 18. P. 145–180.
85. Skulachev V. P. et al. 2017. Neoteny, prolongation of youth: from naked mole rats to «naked apes» (humans) // Physiol Rev. V. 97. P. 699–720.
86. Sohail M. et al. 2017. Negative selection in humans and fruit flies involves synergistic epistasis // Science. 2017. V. 356. P. 539–542.
87. Somel M. et al. 2009. Transcriptional neoteny in the human brain // Proc Natl Acad Sci USA. V. 106. P. 5743–5748.
88. Spaans S. K. et al. 2015. The chromosome copy number of the hyperthermophilic archaeon Thermococcus kodakarensis KOD 1 // Extremophiles. V. 19. P. 741–750.
89. Starita L. M. et al. 2015. Massively parallel functional analysis of BRCA 1 RING domain variants // Genetics. V. 200. P. 413–422.
90. Tan C. K. W. et al. 2013. Sex‑specific responses to sexual familiarity, and the role of olfaction in Drosophila // Proc Biol Sci. V. 280. P. 20131691.
91. Tilszer M. et al. 2006. Evolution under relaxed sexual conflict in the bulb mite Rhizoglyphus robini // Evolution. V. 60. P. 1868–1873.
92. Turkheimer E. et al. 2003. Socioeconomic status modifies heritability of IQ in young children // Psychol Sci. V. 14. P. 623–628.
93. Van’t Hof A. E. et al. 2016. The industrial melanism mutation in British peppered moths is a transposable element // Nature. V. 534. P. 102–105.
94. Venn O. et al. 2014. Strong male bias drives germline mutation in chimpanzees // Science. V. 344. P. 1272–1275.
95. Vlad D. et al. 2014. Leaf shape evolution through duplication, regulatory diversification, and loss of a homeobox gene // Science. V. 343. P. 780–783.
96. Watanabe S. et al. 2016. Color polymorphism in an aphid is maintained by attending ants // Sci Adv. V. 2. P. e1600606.
97. Weekes‑Shackelford V. A., Shackelford T. K. 2004. Methods of filicide: stepparents and genetic parents kill differently // Violence Vict. V. 19. P. 75–81.
98. Wilkins A. S., Holliday R. 2009. The evolution of meiosis from mitosis // Genetics. V. 181. P. 3–12.
99. Williams G. C. 1957. Pleiotropy, natural selection, and the evolution of senescence // Evolution. V. 11. P. 398–411.
100. Wiser M. J. et al. 2013. Long‑term dynamics of adaptation in asexual populations // Science. V. 342. P. 1364–1367.
101. Woodley of Menie M. A. 2015. How fragile is our intellect? Estimating losses in general intelligence due to both selection and mutation accumulation // Pers Indiv Differ. V. 75. P. 80–84.
102. Zeh D. W., Zeh J. A. 2000. Reproductive mode and speciation: the viviparity‑driven conflict hypothesis // Bioessays. V. 22. P. 938–946.
Словарь терминов
Этот словарик призван помочь читателю продраться через сложные разделы книги, не теряя душевного равновесия. Он совершенно не претендует на полноту и строгость определений. Это не энциклопедия, а подсказки, помогающие уловить смысл термина в общих чертах. Дополнительную информацию легко найти в интернете. В словарик не включены базовые понятия, которые, мы надеемся, известны сегодня любому старшекласснику, такие как «геном», «хромосома», «аминокислота», «нуклеотид», «фермент», «популяция» или «экологическая ниша».
Адаптация – процесс роста приспособленности под действием отбора или признак, повышающий приспособленность. Например, можно сказать «Густая шерсть – это адаптация к холодному климату» или «В ходе адаптации к холодному климату многие млекопитающие обзавелись густой шерстью».
Адаптивная радиация – быстрая дивергенция, сопряженная с заполнением разнообразных экологических ниш и с появлением многочисленных новых таксонов. Пример адаптивной радиации – происхождение множества новых видов рыб от немногочисленных особей, случайно попавших в новообразовавшееся озеро.
Аллели‑мутаторы – аллели, повышающие темп мутагенеза. К появлению аллеля‑мутатора может привести, например, мутация, нарушающая работу какого‑нибудь белка, участвующего в репликации или репарации ДНК.
Аллель – вариант гена. Каждый ген в генофонде популяции может присутствовать в виде одного или нескольких аллелей.
Аллопатрия – проживание на разных территориях. Например, два вида называют аллопатрическими, если их ареалы не пересекаются. Если ареалы пересекаются частично, то их перекрывающиеся части называют симпатрическими, а неперекрывающиеся – аллопатрическими.
Археи (или архебактерии) – одна из двух групп прокариот (вторая – бактерии). Археи сильно отличаются от бактерий на молекулярном уровне, хотя внешне выглядят примерно так же. К чести архей нужно заметить, что среди них нет возбудителей инфекционных заболеваний. Возможно, это связано с тем, что археи – не большие мастера по утилизации сложной органики. Даже живя у нас в кишечнике, среди райского изобилия готовых питательных веществ, они синтезируют всю органику для себя сами, используя производимый бактериями молекулярный водород и выделяя в качестве побочного продукта метан. Одна из групп архей некогда дала начало эукариотам.
Аутосомы – все хромосомы, кроме половых. У млекопитающих, например, половыми являются хромосомы X и Y, все прочие – аутосомы.
Бутылочное горлышко – временное снижение численности популяции, после которого численность снова увеличивается.
Вред – на эволюционно‑биологическом жаргоне это слово обычно означает снижение приспособленности. Мутации, снижающие приспособленность, называют вредными.
Гаплоидный – имеющий по одной копии генома в клетках. Когда речь идет о прокариотах, вместо «гаплоидный» принято говорить «моноплоидный».
Генетический автостоп – распространение каких‑либо аллелей под действием отбора, действующего не непосредственно на них, а на другой, сцепленный с ними аллель. Особенно сильно выражен генетический автостоп у одноклеточных, размножающихся клонально (делением, без полового процесса). В такой ситуации отбор может работать только с целыми геномами, но не с отдельными генами, и поэтому полезная мутация может распространиться в генофонде только вместе со всем своим «генетическим фоном», то есть со всеми аллелями, которые имелись в том геноме, где эта мутация возникла. В этом случае только сама полезная мутация распространяется «по‑честному», поскольку повышает приспособленность и поддерживается отбором, а все остальные аллели, распространяющиеся вместе с ней, делают это за счет генетического автостопа.
Генетический груз (мутационный груз) – снижение приспособленности из‑за имеющихся в геноме вредных аллелей (по сравнению с «идеальным» геномом, свободным от таких аллелей).
Генетический дрейф (дрейф генов) – случайные колебания частот встречаемости аллелей, не связанные с действием отбора, то есть с влиянием этих аллелей на приспособленность. В маленьких популяциях дрейф сильнее, чем в больших.
Генотип – совокупность всех наследственных свойств организма, определяемых его ДНК. Иногда термин используется в более узком смысле – в приложении только к одному или нескольким генам, аллельное состояние которых интересует исследователя в данный момент.
Гетерогаметный пол – пол, имеющий две разные половые хромосомы. Например, у млекопитающих и дрозофил гетерогаметный пол – мужской (хромосомы XY). У птиц и бабочек – женский (хромосомы ZW).
Гетеродимер – комплекс из двух разных белковых молекул.
Гетерозиготность – наличие разных аллелей данного гена в разных копиях генома, имеющихся у данного организма. Например, если человек получил от матери один, а от отца другой вариант (аллель) какого‑либо гена, то этот человек гетерозиготен по данному гену.
Гистоны – белки, участвующие в упаковке геномной ДНК и в регуляции активности генов. Характерны для эукариот и некоторых архей, что традиционно рассматривалось как важный аргумент в пользу происхождения эукариот от архей.
Гомогаметный пол – пол, имеющий две одинаковые половые хромосомы. Например, у млекопитающих это женский пол (у самок имеются две Х ‑хромосомы), а у птиц – мужской (у самцов две хромосомы Z).
Гомодимер – комплекс из двух одинаковых белковых молекул.
Гомозиготность – наличие одного и того же аллеля данного гена во всех копиях генома, имеющихся у данного организма. У диплоидных организмов, имеющих две копии генома в каждой клетке, гомозиготной по данному гену будет особь, у которой обе копии этого гена идентичны.
Гомологичные – имеющие общее происхождение (если речь идет, например, о гомологичных органах, таких как рука человека и грудной плавник карася, или белках, таких как человеческий и мышиный гемоглобин) или очень похожую нуклеотидную последовательность (если речь идет о гомологичных хромосомах, способных обмениваться участками при кроссинговере).
Движущий отбор – преимущественное выживание и размножение особей, у которых состояние какого‑то фенотипического признака отличается от среднего по популяции. Под действием движущего отбора среднее значение признака в популяции меняется (если, конечно, наследуемость признака не равна нулю, но признаков с нулевой наследуемостью почти не бывает).
Делеция – мутация, состоящая в потере (выпадении) участка ДНК любой длины: от одного нуклеотида до крупного фрагмента хромосомы.
Дендритный шипик – маленький отросток нейрона, служащий для приема входящих сигналов от другого нейрона. Новые дендритные шипики образуются в ходе обучения, а исчезновение дендритных шипиков связано с забыванием.
Дивергенция – разделение, расхождение. Обычно говорят о дивергенции популяций или видов. Это означает, что некая предковая популяция разделилась и между ее частями ослабел или полностью прекратился обмен генами. После этого популяции начинают накапливать отличия друг от друга, что со временем приводит к появлению у них несовместимых аллелей и развитию постзиготической репродуктивной изоляции (пониженной приспособленности гибридов).
Диплоидный – имеющий две копии генома в каждой клетке.
Естественный отбор – избирательное выживание и размножение более приспособленных индивидов, ведущее к росту частоты встречаемости аллелей, повышающих приспособленность.
Зародышевая линия – половые клетки и их предшественники; потенциально бессмертное подмножество клеток, составляющих многоклеточный организм.
Инбредная депрессия – снижение приспособленности потомства при близкородственных скрещиваниях. Основная причина инбредной депрессии – в переходе рецессивных вредных аллелей в гомозиготное состояние.
Инвазивный вид – вид, занесенный в новый для него регион и успешно размножившийся там.
Индикатор приспособленности – признак, по которому можно оценить приспособленность особи. Например, для многих животных неплохими индикаторами приспособленности служат крупный размер, симметричность, яркая окраска, лоснящаяся густая шерсть и т. п. Под действием полового отбора у животных может развиться склонность выбирать себе половых партнеров с учетом индикаторов приспособленности.
Клада – группа, включающая общего предка и всех его потомков; отдельная целая веточка эволюционного дерева.
Клон – совокупность генетически идентичных особей или потомков одного организма, размножающегося бесполым путем, например почкованием или делением пополам.
Клональная интерференция – конкуренция между клонами в бесполой популяции. Например, если у двух бактерий возникнут две разные полезные мутации, то при отсутствии полового размножения (или хотя бы интенсивного горизонтального переноса генов) эти мутации не смогут объединиться в одном геноме. Вместо этого потомки двух удачливых мутантов будут конкурировать друг с другом, пока один из клонов не будет вытеснен другим. В результате из двух полезных мутаций зафиксируется в генофонде только одна, а вторая будет потеряна. При половом размножении зафиксируются, скорее всего, обе мутации.
Комбинативная изменчивость – разнообразие генотипов, возникающее из‑за перемешивания частей геномов разных особей при половом процессе. Менее эффективным, но все же работающим способом создания комбинативной изменчивости является горизонтальный перенос генов.
Конфликт полов – метафорический термин, описывающий ситуацию, когда под действием полового отбора у одного из полов развиваются адаптации, выгодные этому полу, но вредные для другого. Склонность самок богомолов съедать своих партнеров во время спаривания или склонность самцов многих млекопитающих убивать детенышей, рожденных самками не от них, – яркие проявления конфликта полов.
Конъюгация (у прокариот) – один из способов горизонтального переноса генов, в ходе которого одна клетка (донор) передает другой (реципиенту) часть своего генетического материала.
Коэволюция – согласованная эволюция представителей двух или более видов. Например, паразиты и их хозяева постоянно адаптируются друг к другу, совершенствуя средства защиты и нападения.
Локус – намеренно расплывчатый генетический термин, обозначающий «какой‑то участок генома». При этом может иметься в виду как большой участок, включающий десятки генов и сотни тысяч нуклеотидов, так и одна конкретная нуклеотидная позиция.
Мейоз – особый вид клеточного деления, характерный для эукариот. В результате мейоза число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой. Мейоз также называют редукционным делением, потому что происходит редукция числа копий генома в клетке (плоидность уменьшается вдвое). В ходе мейоза, как правило, происходит кроссинговер – обмен участками между гомологичными хромосомами.
Метагеномный анализ – метод, состоящий в том, что из какой‑то пробы (будь то почва, морская вода или содержимое чьего‑то кишечника) выделяют ДНК, секвенируют, а потом по отсеквенированным фрагментам пытаются понять, какие виды организмов и в каком количестве присутствовали в пробе.
Митоз – вид клеточного деления, характерный для эукариот. Его особенностью является строго упор








