 2020-09-24
2020-09-24 2242
2242Общий план организации
Вестибулярная сенсорная система состоит из следующих отделов:
периферический отдел включает два образования, содержащие механорецепторы вестибулярной системы — преддверие (мешочек и маточка) и полукружные каналы;
проводниковый отдел начинается от рецепторов волокнами биполярной клетки (первого нейрона) вестибулярного узла, расположенного в височной кости, другие отростки этих нейронов образуют вестибулярный нерв и вместе со слуховым нервом в составе 8-ой пары черепно-мозговых нервов входят в продолговатый мозг; в вестибулярных ядрах продолговатого мозга находятся вторые нейроны, импульсы от которых поступают к третьим нейронам в таламусе (промежуточный мозг);
корковый отдел представляют четвертые нейроны, часть которых представлена в проекционном (первичном) поле вестибулярной системы в височной области коры, а другая часть — находится в непосредственной близости к пирамидным нейронам моторной области коры и в постцентральной извилине. Точная локализация коркового отдела вестибулярной сенсорной системы у человека в настоящее время не установлена.
Строение и функции вестибулярного анализатора
Вестибулярный анализатор имеет важное значение в регуляции положения тела в пространстве и его движений. Периферический отдел вестибулярного анализатора является частью внутреннего уха и состоит из полукружных каналов, размещенных в трех взаимно перпендикулярных плоскостях, и из статоцистных органов – двух мешочков – овального (маточки) и круглого, который расположен ближе к улитке.
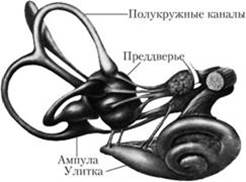
Вестибулярный орган (орган равновесия, орган гравитации) состоит из трех полукружных каналов и преддверия.Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях: верхний—во фронтальной,задний—в сагиттальной инаружный—в горизонтальной. Преддверие состоит из двух мешочков—круглого (саккулюс), расположенного ближе к улитке, иовального (утрикулюс)расположенного ближе к полукружным каналам.
Полукружные каналы своими устьями открываются в преддверие и сообщаются с ним пятью отверстиями (колена двух каналов: верхнего и заднего — соединены вместе). Один конец каждого канала имеет расширение, которое называется ампулой. Все эти структуры состоят из тонких перепонок и образуют перепончатый лабиринт, внутри которого находится эндолимфа.Вокруг перепончатого лабиринта и между ним и костным его футляром имеетсяперилимфа,которая переходит в перилимфу органа слуха. В каждом мешочке преддверия имеются небольшие возвышения, называемые пятнами, а в ампулах полукружных каналов — гребешками. Они состоят из нейроэпителиальных клеток, имеющих на свободной поверхности волоски (реснички), которые разделяются на две группы: тонкие (их много) — стереоцилии и один более толстый и длинный на периферии пучка—киноцилия
Волосковые клеткипредставляют собой рецепторы вестибулярного анализатора иявляются вторичными. Рецепторные клетки преддверия покрыты желеобразной массой, состоящей в основном из мукополисахаридов; благодаря содержанию в ней значительного количества кристаллов карбоната кальция она получила названиеотолитовой мембраны. В ампулах полукружных каналов желеобразная масса не содержит солей кальция и называетсялистовидной мембраной (купула). Волоски рецепторных клеток пронизывают эти мембраны.
Возбуждение волосковых клеток происходит вследствие скольжения мембраны по волоскам, изгибания волосков (стереоцилии) в сторону киноцилий. Это связано с тем, что механическое управление ионными каналами мембраны волоска с помощью микрофиламентов, зависит от направления сгиба волоска:отклонение в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении вызывает закрытие каналов и гиперполяризацию рецептора.
В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва.
проводниковый отдел. К рецепторам подходят периферическиеволокна биполярных нейронов вестибулярного ганглия, расположенного во вутреннем слуховом проходе (первый нейрон). Аксоны этих нейронов в составе вестибулярного нерва направляются квестибулярым ядрам продолговатогомозга (второй нейрон). Вестибулярные ядра продолговатого мозга (верхнее—ядро Бехтерева,медиальное—ядро Швальбе,латеральное—ядро Дейтерсаинижнее—ядро Роллера) получают дополнительную информацию по аф- ферентным нейронам от проприорецепторов; мышц или от суставных сочленений шейного отдела позвоночника. Эти ядра, где расположен второй нейрон вестибулярного анализатора, тесносвязаны с различными отделами центральной нервной системы.Благодаря этому обеспечиваются контроль и управление эффекторными реакциями соматического, вегетативного и сенсорного характера.
третий нейронрасположен вядрах зрительного бугра, откуда возбуждение направляется в кору большого полушария.
Центральный отдел вестибулярного анализатора локализуется ввисочной области коры большого мозга, несколько кпереди от слуховой проекционной зоны (21—22-е поля по Бродману, четвертый нейрон).
Нервные волокна, выходящие из вестибулярных ядер, образуют связи с другими отделами центральной нервной важнейшими из них являются следующие: вестибулоспинальные, вестибуловегетативныеивестибулоглазодвигательные. Эти связи обеспечивают контроль и управление различными двигательными реакциями, а также являются основой для рефлексов обеспечивающих равновесие.
а. Вестибулоспинальный тракт, волокна которого в конечном итоге оказывают влияние в основном наγ-мотонейроны мышц-разгибателей, хотя часть волокон оканчивается и наα-мотонейронах.
б. Связи с мотонейронамишейного отдела спинного мозга, входящие главным образом в вестибулоспинальный тракт.
в.Связи с ядрамиглазодвигательного нерва, которые опосредуют движения глаз, вызываемые вестибулярной активностью; эти волокна проходят в составе медиального продольного пучка. Функцию статолитового аппарата можно тестировать, наблюдая глазодвигательные реакции при наклонах головы.
г. Тракты, направляющиеся ввестибулярные ядра противоположной стороны мозга, благодаря которым афферентация с обеих сторон тела может обрабатываться совместно.
д. Связи сретикулярной формацией, посредством которых обеспечивается воздействие на ретикулоспинальный тракт, являющийся еще одним (полисинаптическим) путем кα- иβ-мотонейронам.
е. Тракты, проходящие через таламус впостцентральную извилину коры головного мозга, обеспечивающие сознательную обработку вестибулярной информации и таким образом сознательную ориентацию в пространстве.
ж.Волокна, направляющиеся вгипоталамус, которые в основном участвуют в возникновении кинетозов (укачивания).
з. Связи смозжечком, в особенности сarchicerebellum(вторичные вестибулярные афференты).мозжечок, может получать некоторые первичные вестибулярные афференты (так называемый прямой сенсорный мозжечковый путь). И первичные, и вторичные вестибулярные афференты у млекопитающих оканчиваются в мозжечке мшистыми волокнами на клетках-зернах флокку-лонодулярной доли (относящейся кarchicerebellum) и частично клеткахuvulaиparaflocculus(paleocerebellum). Клетки-зерна оказывают возбуждающее действие на клетки Пуркине в этих областях, а аксоны последних проецируются опять-таки в вестибулярные ядра. Такая цепь осуществляет тонкую регулировку вестибулярных рефлексов. При нарушениях функции мозжечка вследствие различных заболеваний эти рефлексы утрачивают тормозной компонент, что проявляется в возникновении таких симптомов, как усиленный или спонтанно возникающий нистагм, утрата равновесия, проявляющаяся в неустойчивой походке, избыточной амплитуде движений, особенно при ходьбе («петушиный шаг»). Эти симптомы являются частью синдромамозжечковой атаксии.








