 2020-09-26
2020-09-26 518
518Таламусы (thalami), или зрительные бугры, расположены по бокам III желудочка и составляют до 80% массы промежуточного мозга. Они имеют яйцевидную форму, приблизительный объем 3,3 куб. см и состоят из клеточных скоплений (ядер) и прослоек белого вещества. В каждом таламусе различают четыре поверхности: внутреннюю, наружную, верхнюю и нижнюю.
Внутренняя поверхность таламуса образует боковую стенку III желудочка. От расположенного ниже подбугорья она отделена неглубокой гипоталами-ческой бороздой (sulcus hypothalamics), идущей от межжелудочкового отверстия к входу в водопровод мозга. Внутреннюю и верхнюю поверхности разграничивает мозговая полоска {stria medullaris thalami). Верхняя поверхность таламуса, как и внутренняя, свободна. Она прикрыта сводом и мозолистым телом, с которыми не имеет сращений. В передней части верхней поверхности таламуса расположен его передний бугорок, который иногда называют возвышением переднего ядра. Задний конец таламуса утолщен — это так называемая подушка таламуса (pulvinar). Наружный край верхней поверхности таламуса подходит к хвостатому ядру, от которого ее отделяет пограничная полоска (stria terminalis).
По верхней поверхности таламуса в косом направлении проходит сосудистая борозда, которую занимает сосудистое сплетение бокового желудочка. Эта борозда делит верхнюю поверхность таламуса на наружную и внутреннюю части. Наружная часть верхней поверхности таламуса покрыта так называемой прикрепленной пластинкой, составляющей дно центрального отдела бокового желудочка мозга.
Наружная поверхность таламуса прилежит к внутренней капсуле, отделяющей ее от чечевичного ядра и головки хвостатого ядра. За подушкой таламуса расположены коленчатые тела, относящиеся к метаталамусу. Остальная часть нижней стороны таламуса сращена с образованиями гипоталамической области.
Таламусы находятся на пути восходящих трактов, идущих от спинного мозга и ствола мозга к коре больших полушарий. Они имеют многочисленные связи с подкорковыми узлами, проходящими главным образом через петлю чечевичного ядра (ansa lenticularis). В состав таламуса входят клеточные скопления (ядра), отграниченные друг от друга прослойками белого вещества. К каждому ядру подходят собственные афферентные и эфферентные связи. Соседние ядра формируют группы. Выделяют: I) передние ядра (лис//, anteriores) — имеют реципрокные связи с сосцевидным телом и сводом, известные как сосцевидно-таламический пучок (пучок Вик д'Азира) с поясной извилиной, относящиеся к лимбической системе; 2) задние ядра, или ядра подушки бугра (nucli posteriores) — связаны с ассоциативными полями теменной и затылочной областей; играют важную роль в интеграции различных видов поступающей сюда сенсорной информации; 3) дорсальное боковое ядро (nucl. dorsolateral) — получает афферентные импульсы от бледного шара и проецирует их в каудальные отделы поясной извилины; 4) вентролатералъные ядра (nucli ventrolaterales) — самые крупные специфические ядра, являются коллектором большинства соматосенсорных путей: медиальная петля, спиноталамические пути, тройнично-таламические и вкусовые пути, по которым проходят импульсы глубокой и поверхностной чувствительности и др.; отсюда нервные импульсы направляются в корковую проекционную соматосенсорную зону коры (поля 1, 2, За и 36, по Бродману); 5) медиальные ядра (nucli mediates) — ассоциативные, получают афферентные импульсы от вентральных и интраламинарных таламических ядер, гипоталамуса, ядер среднего мозга и бледного шара; эфферентные пути отсюда направляются в ассоциативные области префронтальной коры, расположенные впереди
моторной зоны; 6) внутрипластинчатые ядра (интраламинарные ядра, nucll. intralaminares) — составляют основную часть неспецифической проекционной системы таламуса; афферентные импульсы они получают частично по восходящим волокнам ретикулярной формации ствола нерва, частично по волокнам, начинающимся от ядер таламуса. Исходящие от этих ядер проводящие пути направляются в хвостатое ядро, скорлупу, бледный шар, относящиеся к экстрапирамидной системе, и, вероятно, в другие ядерные комплексы таламуса, которые затем направляют их во вторичные ассоциативные зоны коры мозга. Важной частью интраламинарного комплекса является центральное ядро таламуса, представляющее таламический отдел восходящей ретикулярной активирующей системы.
Таламусы являются своеобразным коллектором чувствительных путей, местом, в котором концентрируются все пути, проводящие чувствительные импульсы, идущие от противоположной половины тела. Кроме того, в переднее его ядро по сосцевидно-таламическому пучку поступают обонятельные импульсы; вкусовые волокна (аксоны вторых нейронов, расположенных в одиночном ядре) заканчиваются в одном из ядер вентролатеральной группы.
Таламические ядра, получающие импульсы от строго определенных участков тела и передающие эти импульсы в соответствующие ограниченные зоны коры (первичные проекционные зоны), называются проекционными, специфическими или переключающими ядрами. К ним относятся вентролатеральные ядра. Переключающие ядра для зрительных и слуховых импульсов заложены соответственно в латеральных и медиальных коленчатых телах, прилежащих к задней поверхности зрительных бугров и составляющих основную массу забу-горья.
Наличие в проекционных ядрах таламуса, прежде всего в вентролатеральных ядрах, определенного соматотопического представительства делает возможным при ограниченном по объему патологическом очаге в таламусе развитие расстройства чувствительности и сопряженных с этим двигательных нарушений в какой-либо ограниченной части противоположной половины тела.
Ассоциативные ядра, получая чувствительные импульсы от переключающих ядер, подвергают их частичному обобщению — синтезу; в результате из этих таламичсских ядер к коре большого мозга направляются импульсы, уже усложненные вследствие синтеза поступающей сюда информации. Следовательно, таламусы являются не только промежуточным центром переключения, но могут быть и местом частичной переработки чувствительных импульсов.
Кроме переключающих и ассоциативных ядер, в таламусах находятся, как уже упоминалось, интраламинарные (парафасцикулярное, срединное и медиальное, центральные, парацентральное ядра) // ретикулярные ядра, не имеющие специфической функции. Они рассматриваются как часть ретикулярной формации и объединяются под названием неспецифической дидЬфузной тала-мической системы. Будучи связанной с корой больших полушарий и структурами лимбико-ретикулярного комплекса. Эта система принимает участие в регуляции тонуса и в «настройке» коры и играет определенную роль в сложном механизме формирования эмоций и соответствующих им выразительных непроизвольных движений, мимики, плача и смеха.
Таким образом, к таламусам по афферентным путям сходится информация практически от всех рецепторных зон. Эта информация подвергается существенной переработке. Отсюда к коре больших полушарий направляется лишь
часть ее, другая же и, вероятно, большая часть принимает участие в формировании безусловных и, возможно, некоторых условных рефлексов, дуги которых замыкаются на уровне таламусов и образований стриопаллидарной системы. Таламусы являются важнейшим звеном афферентной части рефлекторных дуг, обусловливающих инстинктивные и автоматизированные двигательные акты, в частности привычные локомоторные движения (ходьба, бег, плавание, езда на велосипеде, катание на коньках и т.п.).
Волокна, идущие от таламуса к коре больших полушарий мозга, принимают участие в формировании заднего бедра внутренней капсулы и лучистого венца и образуют так называемые лучистости таламуса — переднюю, среднюю (верхнюю) и заднюю. Передняя лучистость связывает переднее и отчасти внутреннее и наружное ядра с корой лобной доли. Средняя лучистость таламуса — самая широкая — связывает вентролатеральные и медиальные ядра с задними отделами лобной доли, с теменной и височной долями мозга. Задняя лучистость состоит главным образом из зрительных волокон (radiatio optica, или пучок Грациоле), идущих от подкорковых зрительных центров в затылочную долю, к корковому концу зрительного анализатора, расположенному в области шпорной борозды (fissura calcarina). В составе лучистого венца проходят и волокна, несущие импульсы от коры больших полушарий к таламусу (корково-таламические связи).
Сложность организации и многообразие функций таламуса определяет полиморфизм возможных клинических проявлений его поражения. Поражение вен-тролатеральной части таламуса обычно ведет к повышению порога чувствительности на стороне, противоположной патологическому очагу, при этом меняется аффективная окраска болевых и температурных ощущений. Больной воспринимает их как трудно локализуемые, разлитые, имеющие неприятный, жгучий оттенок. Характерна в соответствующей части противоположной половины тела гипалгезия в сочетании с гиперпатией, при этом особенно выражено расстройство глубокой чувствительности, что может вести к неловкости движений, сенситивной атаксии.
При поражении заднелатеральной части таламуса может проявиться так называемый таламический синдром Дежерина—Русси (описали в 1906 г. французские невропатологи J. Dejerine (1849—1917) и G. Roussy (1874—1948)1, включающий в себя жгучие, мучительные, подчас невыносимые таламинеские боли в противоположной половине тела в сочетании с нарушением поверхностной и особенно глубокой чувствительности, псевдоастериогнозом и сенситивной ге-миатаксией, явлениями гиперпатии и дизестезии. Таламический синдром Дежерина—Русси чаще возникает при развитии в нем инфарктного очага в связи с развитием ишемии в латеральных артериях таламуса (аа. thalamki iaterales) — ветвях задней мозговой артерии. Иногда при этом на стороне, противоположной патологическому очагу, возникает преходящий гемипарез и развивается гомонимная гемианопсия. Следствием расстройства глубокой чувствительности может быть сенситивная гемиатаксия, псевдоастриогноз. В случае поражения медиальной части таламуса, зубчато-таламического пути, по которому к таламусу проходят импульсы от мозжечка, и руброталамических связей на противоположной патологическому очагу стороне появляется атаксия в сочетании с атетоидным или хореоатетоидным гиперкинезом, обычно особенно выраженным в кисти и пальцах («таламическая» рука). В таких случаях характерна тенденция к фиксации руки в определенной позе: плечо прижато к туловищу, предплечье и кисть согнуты и пронированы, основные фаланги пальцев
согнуты, остальные разогнуты. Пальцы руки при этом совершают медленные вычурные движения атетоидного характера.
В артериальном кровоснабжении таламуса участвуют задняя мозговая артерия, задняя соединительная артерия, передняя и задние ворсинчатые артерии.
27. Эпиталамус. Эпиталамус (epithalamus, надбугорье) можно рассматривать как непосредственное продолжение крыши среднего мозга. К эниталамусу принято относить заднюю эпиталамическую спайку (commissura epithalamica posterior), два поводка (habenulae) и их спайку {commissura habenularum), а также шишковидное тело (corpus pineale, эпифиз).
Эпиталамическая спайка располагается над верхней частью водопровода мозга и представляет собой комиссуральный пучок нервных волокон, который берет начало от ядер Даркшевича и Кахаля. Впереди от этой спайки расположено непарное шишковидное тело, имеющее вариабельные размеры (при этом длина его не превышает 10 мм) и форму конуса, обращенного вершиной назад. Основание шишковидного тела образовано нижней и верхней мозговыми пластинками, которые окаймляют выворот шишковидного тела (recessus pinealis) — выступающую верхнезаднюю часть третьего желудочка мозга. Нижняя мозговая пластинка продолжается назад и переходит в эпиталамическую спайку и пластинку четверохолмия. Передняя часть верхней мозговой пластинки переходит в спайку поводков, от конца которой отходят направляющиеся вперед поводки, называемые иногда ножками шишковидного тела. Каждый из поводков тянется к зрительному бугру и на границе верхней и внутренней его поверхности заканчивается треугольным расширением, находящимся над расположенным уже в веществе таламуса небольшим ядром уздечки. От ядра уздечки вдоль задненаружной поверхности таламуса тянется белая полоска — stria medullaris, состоящая из волокон, соединяющих шишковидное тело со структурами обонятельного анализатора. В связи с этим существует мнение о том, что эпиталамус имеет отношение к обонянию.
В последнее время установлено, что отделы эпиталамуса, главным образом шишковидное тело, продуцируют физиологически активные вещества — серо-тонин, мелатонин, адреногломерулотропин и антигипоталамический фактор.
Шишковидное тело представляет собой железу внутренней секреции. Оно имеет дольчатое строение, паренхима его состоит из пинеоцитов, эпителиаль-
ных и глиальных клеток. Шишковидное тело содержит большое количество кровеносных сосудов, кровоснабжение его обеспечивается ветвями задних мозговых артерий. Подтверждает эндокринную функцию эпифиза и его высокая способность к поглощению радиоактивных изотопов 32Р и 13Ч. Он поглошает радиоактивного фосфора больше, чем любой другой орган, а по количеству поглощаемого радиоактивного йода уступает только щитовидной железе. До периода полового созревания клетки шишковидного тела выделяют вещества, тормозящие действие гонадотропного гормона гипофиза, и в связи с этим задерживают развитие половой сферы. Это подтверждают клинические наблюдения преждевременного полового созревания при заболеваниях (главным образом при опухолях) шишковидного тела. Существует мнение, что шишковидное тело находится в состоянии антагонистической корреляции со щитовидной железой и надпочечниками и влияет на обменные процессы, в частности на витаминный баланс и функцию вегетативной нервной системы.
Некоторое практическое значение имеет наблюдаемое после полового созревания отложение в шишковидном теле солей кальция. В связи с этим на краниограммах взрослых людей видна тень обызвествленного шишковидного тела, которое при объемных патологических процессах (опухоль, абсцесс и т.п.) в полости супратенториального пространства может смещаться в сторону, противоположную патологическому процессу.
28. Гипоталамус: классификация ядер и связи. Гипоталамус (hypothalamus) составляет нижнюю, филогенетически наиболее древнюю часть промежуточного мозга. Условная граница между таламусами и гипоталамусом проходит на уровне гипоталамических борозд, находящихся на боковых стенках третьего желудочка мозга.
Гипоталамус (рис. 12.4) условно делится на две части: переднюю и заднюю. К задней части гипоталамической зоны относят расположенные позади серого бугра сосцевидные тела (corpora mammillaria) с прилежащими к ним участками мозговой ткани. К передней части относится зрительный перекрест (chiasma opticum) и зрительные тракты (tracti optici), серый бугор (tuber cinereum), воронка (infundibulum) и гипофиз (hypophysis). Гипофиз, соединенный с серым бугром через воронку и гипофизарную ножку, располагается в центре основания черепа в костном ложе — гипофизарной ямке турецкого седла основной кости. Диаметр гипофиза составляет не более 15 мм, масса его от 0,5 до 1 г.
Гипоталамическая область состоит из многочисленных клеточных скоплений — ядер и пучков нервных волокон. Основные ядра гипоталамуса можно разделить на 4 группы.
1. В переднюю группу входят медиальные и латеральные предоптическое, супраоптическое, паравентрикулярные и переднее гипоталамическое ядра.
2. Промежуточную группу составляют дугообразное ядро, серобугорные ядра, вентромедиальное и дорсомедиальнос гипоталамические ядра, дорсальное гипоталамическое ядро, заднее паравентрикулярное ядро, ядро воронки.
3. Задняя группа ядер включает заднее гипоталамическое ядро, а также медиальные и латеральные ядра сосцевидного тела.
4. К дорсальной группе относятся ядра чечевицеобразной петли.
Ядра гипоталамуса имеют ассоциативные связи между собой и с другими отделами мозга, в частности с лобными долями, лимбическими структура 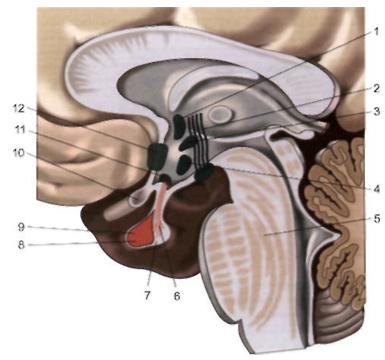 ми больших полушарий, различными отделами обонятельного анализатора, таламусами, образованиями экстрапирамидной системы, ретикулярной формацией ствола мозга, ядрами черепных нервов. Большинство этих связей — двусторонние. Ядра гипоталамической области связывают с гипофизом проходящий через воронку серого бугра и ее продолжение — гипофизарную ножку — гипоталамо-гипофизарный пучок нервных волокон и густая сеть сосудов.
ми больших полушарий, различными отделами обонятельного анализатора, таламусами, образованиями экстрапирамидной системы, ретикулярной формацией ствола мозга, ядрами черепных нервов. Большинство этих связей — двусторонние. Ядра гипоталамической области связывают с гипофизом проходящий через воронку серого бугра и ее продолжение — гипофизарную ножку — гипоталамо-гипофизарный пучок нервных волокон и густая сеть сосудов.
Рис. 12.4. Сагиттальный срез гипоталамуса.
1 — паравентрикулярное ядро; 2 — сосцевидно-таламический пучок; 3 — дорсомеди-альное гипоталамическое ядро; 4 — вентромедиальное гипоталамическос ядро, 5 — мост мозга; 6 — супраоптический гипофизарный путь; 7 — нейрогипофиз; 8 — аде-ногипофиз; 9 — гипофиз; 10 — зрительный перекрест; 11 — супраоптичсское ядро; 12 — преоптическое ядро.
29. Гипофиз: доли гипофиза, особенности связи с гипоталамусом. Гипофиз (hypophisis) представляет собой неоднородное образование. Он развивается из двух разных зачатков. Передняя, большая, его доля (аденогипофиз) формируется из эпителия первичной ротовой полости или так называемого кармана Ратке; она имеет железистое строение. Задняя доля состоит из нервной ткани (нейрогипофиз) и представляет собой непосредственное продолжение воронки серого бугра. Кроме передней и задней долей, в гипофизе различают среднюю, или промежуточную, долю, представляющую собой узкую эпителиальную прослойку, содержащую пузырьки (фолликулы), наполненные серозной или коллоидной жидкостью.
По функции структуры гипоталамуса делят на неспецифические и специфические. Специфические ядра обладают способностью выделять химические
соединения, обладающие эндокринной функцией, регулирующие, в частности, метаболические процессы в организме и поддержание гомеостаза. К специфическим относят обладающие способностью к нейрокринии супраопти-ческое и паравентрикулярное ядра, связанные с нейрогипофизом с помощью супраоптико-гипофизарного пути. Они продуцируют гормоны вазопрессин и окситоцин, которые по упомянутому пути переносятся через ножку гипофиза в нейрогипофиз.
Вазопрессин, или антидиуретический гормон (АДГ), продуцируемый главным образом клетками супраоптического ядра, очень чувствителен к изменению солевого состава крови и регулирует водный метаболизм, стимулируя резорбцию воды в дистальном отделе нефронов. Таким образом, АДГ регулирует концентрацию мочи. При дефиците этого гормона в связи с поражением упомянутых ядер увеличивается количество выделяемой мочи с низкой относительной плотностью — развивается несахарный диабет, при котором наряду с полиурией (до 5 л мочи и более) возникает сильная жажда, ведущая к потреблению большого количества жидкости (полидипсия).
Окситоцин продуцируется паравентрикулярными ядрами, он обеспечивает сокращения беременной матки и влияет на секреторную функцию молочных желез.
Кроме того, в специфических ядрах гипоталамуса образуются «освобождающие» факторы (рилизинг-факторы) и «ингибирующие» факторы, поступающие из гипоталамуса в переднюю долю гипофиза по бугорно-гипофизарному пути (tractus tuberoinfundibularis) и портальной сосудистой сети гипофизарной ножки. Попадая в гипофиз, указанные факторы регулируют секрецию гормонов, выделяемых железистыми клетками передней доли гипофиза.
Клетки аденогипофиза, продуцирующие гормоны под влиянием поступающих в него рилизинг-факторов, являются крупными и хорошо окрашивающимися (хромофильными), при этом большая часть из них окрашивается кислыми красками, в частности эозином. Их называют эозинофильными, или оксифильными, а также альфа-клетками. Они составляют 30—35% всех клеток аденогипофиза и продуцируют соматотропный гормон (СТГ)* или гормон роста (ГР), а также пролактин (ПРЛ). Клетки аденогипофиза (5—10%), окрашивающиеся щелочными (основными, базисными) красками, в том числе гематоксилином, называются базофильными клетками, или бета-клетками. Они выделяют адренокортикотропный гормон (АКТГ) и тиреотропный гормон (TIT).
Около 60% клеток аденогипофиза плохо воспринимают краски (хромофобные клетки, или гамма-клетки) и не обладают гормоносекреторной функцией.
Источниками кровоснабжения гипоталамуса и гипофиза являются ветви артерий, составляющих артериальный круг большого мозга (circulus arteriosis cerebri, виллизиев круг), в частности гипоталамические ветви средней мозговой и задней соединительной артерий, при этом кровоснабжение гипоталамуса и гипофиза оказывается исключительно обильным. В I мм3 ткани серого вещества гипоталамуса насчитывается в 2—3 раза больше капилляров, чем в таком же объеме ядер черепных нервов. Кровоснабжение гипофиза представлено так называемой воротной (портальной) сосудистой системой. Отходящие от артериального круга артерии разделяются на артериолы, затем образуют густую первичную артериальную сеть. Обилие сосудов гипоталамуса и гипофиза обеспечивает происходящую здесь своеобразную интеграцию функций нервной, эндокринной и гуморальной систем. Сосуды гипоталамической области и гипофиза обладают высокой проницаемостью для различных химических и гормональных
ингредиентов крови, а также белковых соединений, в том числе нуклеопроте-идов, нейротропных вирусов. Это определяет повышенную чувствительность гипоталамической области к воздействию разнообразных вредных факторов, попадающих в сосудистое русло, что необходимо хотя бы для обеспечения скорейшего их выведения из организма с целью поддержания гомеостаза.
Гипофизарные гормоны выделяются в кровяное русло и гематогенным путем, достигая соответствующих мишеней. Существует мнение, что частично они попадают в ликворные пути, прежде всего в III желудочек мозга.
Эндокринные функции гипоталамуса и гипофиза регулируются нервной системой. Продуцируемые в них гормоны можно отнести к лигандам — биологически активным веществам, носителям регулирующей информации. Мишенью для них служат специализированные рецепторы органов и тканей. Поэтому гормоны можно рассматривать как своеобразные медиаторы, которые могут передавать информацию на большие расстояния гематогенным путем. В таких случаях этот путь рассматривают как гуморальное колено сложных рефлекторных дуг, обеспечивающих деятельность отдельных органов и тканей на периферии. Кстати, информация о деятельности этих органов и тканей направляется в структуры центральной нервной системы, в частности гипоталамуса, по нервным афферентным путям, а также гематогенным путем, по которому с периферии в центр передается информация о степени активности различных периферических желез внутренней секреции (процесс обратной афферентации).
Такая трактовка роли гормонов исключает представления об автономности эндокринной системы и подчеркивает взаимосвязь и взаимозависимость эндокринных желез и нервной ткани.
Гипоталамические структуры осуществляют регуляцию функций симпатического и парасимпатического отделов вегетативной нервной системы и поддержание в организме вегетативного баланса, при этом в гипоталамусе могут быть выделены эрготропные и трофические зоны (Hess W., 1881 — 1973).
Эрготропная система активирует физическую и психическую деятельность, обеспечивая включение преимущественно симпатических аппаратов вегетативной нервной системы. Трофотропная система способствует накоплению энергии, пополнению затраченных энергетических ресурсов, обеспечивает процессы парасимпатической направленности: тканевый анаболизм, уменьшение частоты сердечных сокращений, стимуляцию функции пищеварительных желез, снижение мышечного тонуса и пр.
Трофотропные зоны находятся главным образом в передних отделах гипоталамуса, прежде всего в его преоптической зоне, эрготропные — в задних отделах, точнее, в задних ядрах и латеральной зоне, которые В. Гесс назвал динамогенными.
Дифференциация функций различных отделов гипоталамуса имеет функционально-биологическое значение и определяет их участие в осуществлении целостных поведенческих актов.
30. Общий план строения конечного мозга: серое и белое вещество. Большой мозг составляет крышу и стенки конечного мозга и достигает крупных размеров, образуя левое полушарие головного мозга и правое полушарие головного мозга, покрывающие сверху большую часть головного мозга.Полушария большого мозга состоят из коры головного мозга и лежащей под ней центральной массы белого вещества головного мозга. Белое вещество состоит из проводящих путей. Левое и правое полушария соединены широким нервным трактом, который называется мозолистым телом. Каждое полушарие для удобства делят на четыре доли. Конечный мозг (telencephalon), или большой мозг (cerebrum) (рис. 9), представляет собой наиболее массивный отдел головного мозга (85-90% массы всего мозга) и занимает большую часть полости черепа. Конечный мозг состоит из двух полушарий (hemispherium) большого мозга, отделенных друг от друга глубокой продольной щелью (fissura longitudinalis cerebri).
Конечный мозг представлен двумя большими полушариями. Большие полушария — самая большая часть головного мозга человека. В норме полушария относительно симметричны и соединены между собой массивным пучком аксонов (мозолистым телом), по которому происходит передача информации из одного полушария в другое. В состав каждого полушария входят базальные ядра, желудочек, белое вешество и плащ, образованный корой.
В соответствии с филогенезом выделяют обонятельный мозг, базальные ядра и кору полушарий. В состав обонятельного мозга, как наиболее филогенетически древней части, входят: обонятельная доля, парагипокампальная извилина, зубчатая извилина, сводчатая извилина, крючок. В каждом полушарии выделяют четыре доли:
лобная, теменная, височная и затылочная.
В полушарии выделят три поверхности:
нижнюю, медиальную и верхнелатералыгую.
Выделяют также и три полюса (самые выступающие части полушарий):
лобный, затылочный и височный.
В каждом полушарии находится латеральный желудочек, являющийся полостью полушария и заполненный ликвором. В каждом из желудочков выделяют передний рог, расположенный в лобной доле, задний рог, расположенный в затылочной доле, нижний рог в толще височной доли и центральную часть. Боковые желудочки через отверстия сообщаются с полостью III желудочка, куда и оттекает ликвор.
Серое вещество больших полушарий представлено так называемыми базальными ядрами: скоплением нервных клеток в глубине полушарий.
31. Борозды и извилины латеральной поверхности полушарий.
32. Борозды и извилины медиальной поверхности полушарий.
33. Борозды, извилины и структуры нижней поверхности полушарий.
Полушария головного мозга, hemispherium cerebralis, снаружи содержат тонкий слой серого вещества - кору больших полушарий. Различают три поверхности полушарий: (1) верхнелатеральная, наиболее выпуклая, facies superlateralis (hemispherii), (2) медиальная, facies medialis (hemispherii), плоская, обращенная к соседнему полушарию и (3) нижнюю, facies inferior (hemispherii), имеющую сложный рельеф, соответствующий внутреннему основанию черепа. Эти поверхности граничат друг с другом краями: верхним, margo superior, нижнелатеральным, margo inferior, и нижнемедиальным, margo medialis. Участки полушарий, наиболее выступающие кпереди и кзади, называют полюсами: лобный полюс, polus frontalis, затылочный полюс, polus ocdpitalis, и височный полюс, polus temporalis. Рельеф поверхностей полушарий очень сложен в связи с наличием более или менее глубоких борозд головного мозга и расположенных между ними выпуклых возвышений - извилин головного мозга. Как и любые живые структуры, структуры нервной системы являются вероятностными структурами. Отсюда, размеры и координаты (глубина, протяженность, форма и направление) борозд и извилин - вероятностные переменные.
В переднем отделе каждого полушария головного мозга находится лобная доля, lobus frontalis. Она заканчивается спереди лобным полюсом и ограничена снизу латеральной бороздой, sulcus lateralis (сильвиева борозда), а сзади - глубокой центральной бороздой (рис. 124, 125). Центральная борозда, sulcus centralis (роландова борозда), расположена во фронтальной плоскости. Она начинается в верхней части медиальной поверхности полушария большого мозга, рассекает его верхний край, спускается, не прерываясь, по верхнелатеральной поверхности полушария вниз и заканчивается, немного не доходя до латеральной борозды. Спереди от центральной борозды, почти параллельно ей, располагается предцентральная борозда, sulcus precentralis. Последняя заканчивается внизу, не доходя до латеральной борозды. Предцентральная борозда нередко прерывается в средней части и состоит из двух самостоятельных борозд. От предцентральной борозды вперед направляются верхняя и нижняя лобные борозды, suici frontales superior et inferior. Они расположены почти параллельно друг другу и делят верхнелатеральную поверхность лобной доли на извилины. Между центральной бороздой сзади и предцентральной бороздой спереди находится предцентральная извилина, gyrus precentralis (anterior). Выше верхней лобной борозды лежит верхняя лобная извилина, gyrus frontalis superior, занимающая верхнюю часть лобной доли. Между верхней и нижней лобными бороздами расположена средняя лобная извилина, gyrus frontalis medius. Книзу от нижней лобной борозды находится нижняя лобная извилина, gyrus frontalis inferior. В эту извилину снизу вдаются ветви латеральной борозды: восходящая ветвь, ramus ascendens, и передняя ветвь, ramus anterior. Эти ветви делят нижнюю часть лобной доли, нависающую над передней частью латеральной борозды, на три части. Покрышечная часть (лобная покрышка), pars opercularis (operculum frontale), расположена между восходящей ветвью и нижним отделом предцентральной борозды. Эта часть лобной доли получила такое название потому, что она прикрывает лежащую в глубине борозды островковую долю (островок). Треугольная часть, pars triangularis, находится между восходящей ветвью сзади и передней ветвью спереди. Глазничная часть, pars orbitalis, лежит книзу от передней ветви, продолжаясь на нижнюю поверхность лобной доли. В этом месте латеральная борозда расширяется, в связи с чем ее называют латеральной ямкой большого мозга, fossa. lateralis (cerebraiis).
Кзади от центральной борозды находится теменная доля, lobus parietalis. Задней границей этой доли является теменно-затылочная борозда, sulcus parietooccipitalis. Эта борозда располагается на медиальной поверхности полушария, глубоко рассекает верхний край полушария большого мозга и переходит на его верхнелатеральную поверхность. Границей между теменной и затылочной долями на дорсолатеральной поверхности полушария большого мозга служит условная линия - продолжение теменно-затылочной борозды книзу. Нижней границей теменной доли является латеральная борозда (ее задняя ветвь), отделяющая эту долю (ее передние отделы) от височной доли.
В пределах теменной доли выделяют постцентральную борозду, sulcus postcentralis. Она начинается от латеральной борозды внизу и заканчивается вверху, не доходя до верхнего края полушария. Постцентральная борозда лежит позади центральной борозды и почти параллельно ей. Между центральной и постцентральной бороздами располагается постцентральная извилина, gyrus postcentralis (gyrus centralis posterior). Вверху она переходит на медиальную поверхность полушария большого мозга, где соединяется с предцентральной извилиной лобной доли, образуя вместе с нею парацентральную дольку, lobulus paracentralis. На верхнелатеральной поверхности полушария, внизу, постцентральная извилина также переходит в предцентральную извилину, охватывая снизу центральную борозду. От постцентральной борозды кзади отходит внутритеменная борозда, sulcus intraparietalis. Она параллельна верхнему краю полушария. Кверху от внутритеменной борозды находится группа мелких извилин, получивших название верхней теменной дольки, lobulus parietalis superior.
Ниже внутритеменной борозды лежит нижняя теменная долька, lobulus parietalis inferior. В ней выделяют две извилины: надкраевую, gyrus supramarginalis, и угловую, gyrus angularis. Надкраевая извилина охватывает конец латеральной борозды, а угловая - конец верхней височной борозды. Нижняя часть нижней теменной дольки и прилежащие к ней нижние отделы постцентральной извилины вместе с нижней частью предцентральной извилины, нависающие над островковой долей, образуют лобно-теменную покрышку островка, operculum frontoparietale.
Затылочная доля, lobus occlpitalis, располагается позади теменно-затылочной борозды и её условного продолжения на верхнелатеральной поверхности полушария. По сравнению с другими долями она имеет небольшие размеры. Затылочная доля кончается затылочным полюсом, polus occipitalis. Борозды и извилины на верхнелатеральной поверхности затылочной доли очень вариабельны. Наиболее часто и лучше других выражена поперечная затылочная борозда, sulcus occipitalis transversus, которая является как бы продолжением кзади внутритеменной борозды теменной доли.
Височная доля, lobus temporalis, занимает нижнебоковые отделы полушария и отделяется от лобной и теменной долей глубокой латеральной бороздой. Край височной доли, прикрывающий островковую (центральную)долю, получил название височной покрышки, operculum temporale. Передняя часть височной доли образует височный полюс, polus temporalis. На боковой поверхности височной доли видны две борозды - верхняя и нижняя височные, suici temporales superior et inferior, почти параллельные латеральной борозде. Извилины височной доли ориентированы вдоль борозд. Верхняя височная извилина, gyrus temporalis superior, расположена между латеральной бороздой вверху и верхней височной внизу. На верхней поверхности этой извилины, скрытой в глубине латеральной борозды, располагаются две - три короткие поперечные височные извилины, gyri temporales transversi (извилины Гешля), разделенные поперечными височными бороздами, suici temporales transversi. Между верхней и нижней височными бороздами находится средняя височная извилина, gyrus temporalis medius. Нижнелатеральный край височной доли занимает нижняя височная извилина, gyrus temporalis inferior, ограниченная сверху одноименной бороздой. Задний конец этой извилины продолжается в затылочную долю.
Островковая доля (островок), lobus insularis (insula), находится в глубине латеральной борозды. Эту долю можно увидеть, если раздвинуть или удалить прикрывающие островок участки лобной, теменной и височной долей, которые получили наименование покрышки. Глубокая круговая борозда островка, sulcus circularis insulae, отделяет островок от окружающих его отделов мозга. На поверхности островка имеются извилины островка, длинная и короткие, gyri insulae (longus et breves). Между длинной извилиной, находящейся в задней части островка и ориентированной сверху вниз и вперед, и короткими извилинами, занимающими верхнепереднюю часть островка, находится центральная борозда островка, sulcus centralis insule. Нижнепередняя часть островка лишена борозд и имеет небольшое утолщение, получившее название порога островка, limen insulae.
| |||
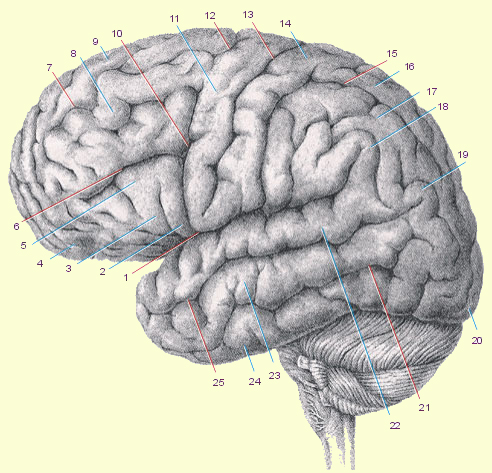
| 8. Средняя лобная извилина, gyrus frontalis medius. 9. Верхняя лобная извилина, gyrus frontalis superior. 10. Нижняя предцентральная борозда, sulcus precentralis inferior. 11. Предцентральная извилина, gyrus precentralis (anterior). 12. Верхняя предцентральная борозда, sulcus precentralis superior. 13. Центральная борозда, sulcus centralis (роландова борозда). 14. Постцентральная извилина, gyrus postcentralis (gyrus centralis posterior). 15. Внутритеменная борозда, sulcus intraparietalis. 16. Верхняя теменная долька, lobulus parietalis superior. 17. Нижняя теменная долька, lobulus parietalis inferior. 18. Надкраевая извилина, gyrus supramarginalis. 19. Угловая извилина, gyrus angularis. 20. Затылочный полюс, polus occipitalis. 21. Нижняя височная борозда, suicus temporalis inferior. 22. Верхняя височная извилина, gyrus temporalis superior. 23. Средняя височная извилина, gyrus temporalis medius. 24. Нижняя височная извилина, gyrus temporalis inferior. 25. Верхняя височная борозда, suicus temporalis superior. | ||








