 2020-10-10
2020-10-10 444
444Введение.
Прежде, чем обсуждать физиологию ЦНС, давайте вспомним, чем представлена ЦНС. Вообще нервная система классифицируется анатомически и функционально. Анатомически нервную систему разделяют на центральную (головной и спинной мозг) и периферическую (черепные, спиномозговые нервы, различные ганглии/узлы). Функционально нервную систему разделяют на соматическую (произвольную, т.е. поддающуюся нашей воле) и вегетативную (непроизвольную, не поддающуюся нашей воле), вегетативная в свою очередь делится на симпатическую (интенсивная деятельность организма) и парасимпатическую (относительный покой и расслабление организма).
Что такое ЦНС, поняли, а каковы её основные функции? Во-первых, центральная-регуляторная – ЦНС регулирует все физиологические функции, при этом не по отдельности, а во взаимосвязи, т.е. ЦНС связует между собой ткани, органы, системы органов между собой и регулирует их для обеспечения нормально функционирования целого организма. Во-вторых, ЦНС обеспечивает связь организма с внешней средой, так как с помощью рецепторов получает всю информацию о внешней среде и регулирует обмен энергией, веществами и информацией между организмом и внешней средой. В-третьих, именно с помощью ЦНС осуществляется наша психическая деятельность.
В разделе регуляции физиологических функций мы говорили о том, что рефлекс лежит в основе нервной регуляции, а сейчас я дополню эти слова, сказав, что рефлекс лежит в основе деятельности всей ЦНС. Рефлекс – это ответная реакция организма на раздражение рецепторов с участием ЦНС. Рефлексы – тема очень обширная, поэтому для них существует ряд классификаций и даже рефлекторная теория. Обсудим это.
Классификация рефлексов. Рефлексы классифицируют по биологическому значению (их много), например, пищевые, оборонительные, половые, позно-тонические и тд. По локализации раздражаемых рецепторов: экстрацептивные (рецепторы внешней среды, расположены в основном в коже, типа температура, давление и тд), интрацептивные (рецепторы внутренней среды, типа давление, концентрация углекислого газа) и пропреоцептивные (расположены в опорно-двигательной системе). По уровню ЦНС, на котором замыкается рефлекторная дуга: спинальные (в спинном мозге), бульбарные (в продолговатом мозге), мезенцефальные (в среднем мозге). По характеру ответной реакции (что делает эффектор): моторные (двигают мышцы), секреторные (вызывают секрецию веществ) и вазомоторные (изменяют просвет сосудов). По приобретённости: условный (образуются в ходе онтогенеза, т.е. вы их сами создаёте) и безусловные (образуются в ходе филогенеза, т.е. вы получаете их по наследству). По морфологическому признаку: соматические и вегетативные. Вот о последних давайте поговорим подробнее.
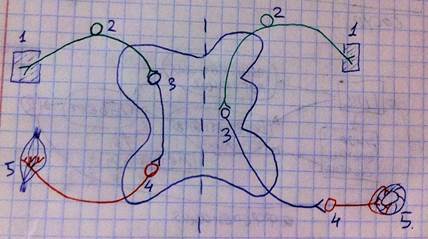
Слева схематично изображена рефлекторная дуга для соматического рефлекса: 1 – рецептор, 2 – афферентный/чувствительный нейрон, 3 – ассоциативный/вставочный нейрон (может быть несколько, а может и не быть), 4 – эфферентный/моторный нейрон, 5 – эффектор. Справа схематично изображена рефлекторная дуга для вегетативного рефлекса: 1 – рецептор, 2 – афферентный/чувствительный нейрон, 3 – преганглионарный нейрон, 4 – постганглионарный нейрон, 5 – эффектор. Как видите, существуют некоторые морфологические отличия.
Теперь о рефлекторной теории. Она была создана И.П. Павловым и И.М. Сеченовым и содержала три основных принципа: принцип детерминизма, принцип структурности, принцип анализа-синтеза. Разберёмся подробно в каждом из них. Принцип детерминизма гласит, что рефлекс не может возникнуть без раздражения рецептора, ибо нейроны сами по себе ПД (и ЛО тоже) не вырабатывают, ПД и ЛО вырабатываются только в рецепторе (участок мембраны с рецепторами), если ничто не раздражает рецептор, то нервный импульс не возникнет. Принцип структурности гласит, что структурной основой рефлекса является рефлекторная дуга (думаю, тут всё ясно). Принцип анализа-синтеза предполагает, что в ходе рефлекса информация, получаемая с рецептора в форме нервного импульса (или скорее импульсов) анализируется, т.е. раскладывается на компоненты, эти компоненты оцениваются, потом оценки складываются (синтез) и принимается решение, какой ответ надо сформировать. Третий принцип станет более понятным, когда мы будем говорить о распространении возбуждения в ЦНС. Это были три принципа классической рефлекторной теории, однако затем П.К. Анохин добавил четвёртое положение об обратной связи, которую мы уже обсуждали в разделе регуляции физиологических функций.
Клетки ЦНС.
Мы обсудили общие вопросы, теперь бы надо приступить к частным, но перед тем, как разбирать функционирование нервной ткани, надо получить базовую информацию об элементах этой ткани – нейронах и глиальных клетках.
Нейроны. Они являются основной структурно-функциональной единицей нервной системы. Почему? ЦНС работает на рефлексах, который предполагают структурную основу в виде рефлекторных дуг, которые состоят из нейронов. Поэтому нейроны – главные клетки в нервной ткани. Разберёмся в их строении.

Нейрон всегда имеет тело (перинуклеон) и отростки (аксон и дендриты, число последних колеблется от нуля до сотен); перед началом аксона формируется аксональный холмик, обладающий повышенной возбудимостью (в синапсах на нейроне возникает множество ВПСП (возбуждающие постсинаптические потенциалы) и ТПСП (тормозящие), которые доходят до аксонального холмика, их воздействие суммируется и если потенциал достаточный, то в аксональном холмике генерируется ПД, который идёт по аксону), ближе к концу аксон может ветвиться на терминалии, которые оканчиваются булавовидными утолщениями, образующими синапсы с другими клетками; на дендритах могут иметься вздутия – шипики. По дендритам сигнал идёт в направлении от отростка к телу, а по аксону от тела к отростку.
Нейроны бывают различными, поэтому нам надо знать их классификацию. Нейроны классифицируются по морфологическому признаку на униполярные (из отростков имеют только аксон), псевдоуниполярные (от тела отходит «отросток», который расходится на дендрит и аксон), биполярные (имеют один дендрит и аксон, не контактирующие друг с другом, как в случае с псевдоуниполярным нейроном), мультиполярные (один аксон, множество дендритов). По функциональному признаку можно разделить на афферентные (имеют рецептор), ассоциативные (не выходят за пределы ЦНС), эфферентные (сообщаются с эффектором), а ещё в функциональном отношении иногда нейроны делят на возбуждающие и тормозные.
Какие функции нейроны выполняют в роли структурно-функциональной единицы ЦНС? Во-первых, интегративная функция. Нейрон, чтобы послать или не послать сигнал на свой аксон, получает импульсы от множества нейронов, он их объединяет (интегрирует все ВПСП и ТПСП) и на их основе решает, отправлять импульс на свой аксон или нет. Во-вторых, координационная функция. Вообще координация – это согласованная последовательная деятельность всех нервных центров при правильной работе ЦНС. Причём же здесь отдельный нейрон? Нервные центры ведь состоят из нейронов, которые должны правильно между собой соединяться и нормально функционировать, именно нейрон координирует сигнал, образовавшийся на нём, путём направления этого сигнала на нужные нейроны, например. В-третьих, трофическая функция. Все электрические явления в нейроне протекают в его отростках и на поверхности тела, отростки надо обеспечивать энергией и нейромедиаторами – этим занимается тело нейрона, выполняя тем самым трофическую функцию для своих отростков.
Второй тип клеток в нервной ткани – глиальные. Это клетки поддержки для нейронов, у них есть много принципиальных разновидностей, поэтому морфологию мы не обсуждаем, а сразу переходим к их функциям: защитная, опорная, изоляционная, обменная, модулирующая. Чуть подробнее. Микроглии обеспечивают иммунитет в нервной ткани. Астроциты фиксируют нейроны в пространстве, как столбики. Те же самые астроциты переносят питательные вещества и кислород из сосуда в нейрон, а из нейрона в сосуд конечные продукты метаболизма (в периферической нервной системе этим занимаются Швановские клетки). Олигодендроциты (а в периферической это опять Швановские клетки) обматывают нейроны, чтобы обеспечить в них изолированное проведение нервных импульсов. И наконец астроциты выделяют в синапс модуляторы, которые изменяют частоту и амплитуду нервных импульсов.
Синапсы.
Мы говорили, что ЦНС работает на основе рефлексов, которые в качестве структурной основы имеют рефлекторную дугу, которая представлена синаптически связанными нейронами. Вот о нейронах мы всё сказали, а вот о способе их связи – синапсах – нам предстоит поговорить. Как вы уже поняли – синапс – межклеточное соединение для нейронов. Какова структура этих соединений? Здесь необходимо отдельно разобрать электрические и химические синапсы.

В электрическом синапсе пресинаптическая мембрана (нейрон, от которого идёт сигнал) и постсинаптическая мембрана (клетка, на которую идёт сигнал) очень сближаются, в них есть белковые каналы-коннексоны, которые присоединяются друг к другу и получается, что у двух клеток сообщается цитоплазма, а сигнал непосредственно проходит с одной клетки на другую. В нашем организме очень мало таких синапсов.
Химические синапсы. Образуются булавовидным утолщением аксона нейрона и какой-нибудь клеткой (нейрон, мышечная, секреторная). В булавовидном утолщении имеются везикулы с нейромедиатором, это же утолщение формирует пресинаптическую мембрану, другая клетка формирует постсинаптическую мембрану, а между ними синаптическая щель. Как происходит сама передача импульса? Посложнее, чем в электрическом синапсе. ПД по аксону доходит до булавовидного утолщения, а ПД это в первую очередь открытие всех натриевых каналов, поэтому я их изобразил на рисунке. Но в булавовидном утолщении есть и каналы для кальция, которые включаются от ПД, впускают кальций в клетку, тот запускает экзоцитоз везикул с нейромедиатором, тот выходит в синаптическую щель и достигает постсинаптической мембраны, действует на её рецепторы и открывает каналы (метаботропные и ионотропные). Если синапс возбуждающий, то открываются каналы для натрия, и мы наблюдаем локальный ответ на мембране (медленная деполяризация, составляющая больше 50%, но меньше 100% от порогового потенциала, т.е. дельты-Е), он же ВПСП. Если синапс тормозной, то открываются каналы для калия (выходит из клетки) и хлора (входит в клетку) – клетка теряет плюсы и приобретает минусы, поэтому потенциал отклоняется в отрицательную сторону, это приводит к истинной гиперполяризации (истинная, а не следовая, потому что происходит самостоятельно, а не как часть ПД), она же ТПСП.

Все ВПСП и ТПСП доходят до аксонального холмика, суммируются и приводят либо не приводят к возбуждению. Но продолжим о механизме передачи сигнала в химических синапсах. Медиатор действует недолго, потому что он: просто рассеивается и уходит из щели, разрушается специальным ферментом, обратно захватывается пресинаптической мембраной, чтобы сигнал не передавался неоправданно долго. Особенность химических синапсов состоит в том, что в одном синапсе может быть только один вид нейромедиатора, а ещё в химических синапсах есть задержка в передаче сигнала (синаптическая задержка), потому что между ПД на пресинаптической мембране и ВПСП/ТПСП на постсинаптической мембране есть ряд действий, отнимающих время: пока кальций наполнит клетку, пока везикулы дойдут до мембраны, пока нейромедиатор доползёт до рецепторов каналов, а если каналы метаботропные, то надо ещё подождать, пока сигнал с рецептора канала передастся на сам канал.
Теперь вы в общих чертах знаете, что такое синапсы, как они устроены и как работают, теперь же обсудим их классификацию. Есть много принципов классификации синапсов. Во-первых, по механизму передачи сигнала: электрические и химические. Во-вторых, по локализации (между какими частями нейронов образован синапс): аксо-дендритные, аксо-шипиковые, аксо-соматические, аксо-аксональные. В-третьих, по выполняемой функции: возбуждающие и тормозящие. В-четвёртых, по клетке, образующей постсинаптическую мембрану: нейро-нейрональный, нервно-мышечный, нервно-эпителиальный.
Нейронные сети.
В ЦНС есть особая организация нейронов, они объединяются не только в рефлекторные дуги, но ещё и в нейронные сети. Нейронная сеть – совокупность синаптически связанных нейронов, участвующая в получении, передаче, хранении, воспроизведении информации. Благодаря нервным сетям возбуждение не идёт по одной протоптанной дорожке, а распрстраняется по ЦНС. Сейчас мы разберём 3 наиболее важные вида сетей (так-то их, конечно, гораздо больше).

Первый тип сетей – дивергентные сети, в которых один нейрон передаёт сигнал на несколько нейронов, сигнал разделяется (амплитуда его не падает, ибо ПД идёт по принципу «всё или ничего»), такая сеть лежит в основе иррадиации – распространения возбуждения по ЦНС в разные нервные центры. Зачем это нужно? Чаще всего информация, поступающая по одному нейрону, представляет интерес для нескольких нервных центров, поэтому приходится делать такую цепь, чтобы распространить сигнал, помимо этого, такая сеть помогает анализировать поступающий сигнал, разбирая его на компоненты.
Второй тип сетей – конвергентные сети, в которых сигналы от нескольких нейронов поступают на один общий и тот решает, посылать сигнал на свой аксон или нет. Такая сеть лежит в основе принципа общего конечного пути. Просто один нейрон, например, может иннервировать конкретный эффектор, за управление которого отвечает несколько нервных центров, нервных центров несколько, а эффектор один – нужна конвергентная сеть, в которой нейрон собирает и суммирует решения нервных центров и сам решает, надо ли генерировать ему ПД или нет. Т.е. дивергентная сеть осуществляет синтез сигналов.
Третий тип сетей – циркулярные, в которых на «основной» нейрон приходит сигнал, он передаёт этот сигнал на следующий нейрон, но ещё он посылает сигнал на дополнительный нейрон, а дополнительный нейрон снова посылает сигнал на основной, тот повторяет действия. Что же происходит? Откуда-то извне в эту сеть пришёл всего один импульс, но он начал циркулировать в этой сети и, проходя каждый цикл, он ещё и отправлялся на следующий нейрон. Итак, на входе в сеть у нас посылается всего один сигнал, а на выходе мы получаем множество. Однако я недаром нарисовал два варианта этой сети: слева примитивный вариант сети, а справа реальный. Особенность реального варианта в том, что добавляется тормозный нейрон, который включается при определённом количестве или частоте циркуляций в сети и выключает эту сеть – без него бы сигнал циркулировал без конца. Данные сети являются основой кратковременной памяти (за счёт циркуляции они могут некоторое время сохранять информацию, полученную даже единожды), ещё они реализуют принципы трансформации сигнала в рефлекторной дуге и рефлекторного последействия, но это мы обсудим позже. Пока что надо запомнить архитектуру цепей, ход сигнала в них и смысл этих сетей. Кстати закорючками на рисунке я обозначил ПД (так делать можно).
Нервные центры.
Продолжаем двигаться по структурам выше: мы обсудили клеточный состав, рефлекторные дуги, нейронные сети, а теперь перейдём к ещё более сложной структуре – нервному центру. Нервный центр – это динамическая совокупность нейронов, координированная деятельность которых обеспечивает регуляцию отдельных функций или определённых рефлекторных актов. Грубо говоря, множество нейронов, синаптически связанных и настроенных на обработку информации и принятие решения по регулированию объекта. Но почему совокупность динамическая? А потому, что при необходимости могут подключаться дополнительные нейроны, это во-первых, а во-вторых, связи между нейронами могут меняться, а сам центр при этом как бы перепрограммируется, проводя обучение/адаптацию. Нервные центры являются основными узловыми компонентами функциональных систем. Исходя из всего сказанного, можно заключить, что нервные центры – это относительно обособленные и функционально значимые элементы нервной системы, поэтому они заслуживают наше внимание. Поговорим об основных свойствах нервных центров, их 6: низкая лабильность, высокая утомляемость, высокая чувствительность к гипоксии, высокая чувствительность к ацидозу и алкалозу, высокая чувствительность к ядам, пластичность.
Низкая лабильность. Насколько вы помните, лабильность – это время, затрачиваемое на разворачивание единичного ПД. Низкая лабильность имеется ввиду, что время-то высокое. Сами нейроны могут обладать высокой лабильностью, но нервный центр – это синаптически связанные нейроны! Т.е. у нас есть нейрон, который в теории может разворачивать ПД за 1 мс, но этого никогда не будет, потому что этот нейрон генерирует ПД, получая сигнал(ы) от другого нейрона(ов), и получает сигналы он через синапс, а «лабильность» в синапсе (обычно понятие лабильность применяют к ПД, а в синапсах ВПСП и ТПСП, поэтому я слово лабильность здесь взял в ковычки) очень низкая вследствие синаптической задержки. Т.е. сам нейрон имеет лабильность 1 мс, а частоту в 1000 Гц, но от других нейронов он получает сигнал только раз в 10 мс (сами ВПСП и ТПСП занимают 0,3 мс, а синаптическая задержка, предшествующая им, займёт 9,7 мс), поэтому лабильность нервного центра в целом 10 мс, а частота 100 Гц. Это был пример, запомните, что причиной низкой лабильности в нервных центрах является синаптическая задержка, ибо нервные центры – это множество синаптически связанных нейронов, причём синапсы в основном химические, а потому и имеют задержку.
Высокая утомляемость. Нервные центры не могут долго и интенсивно работать – их деятельность угасает. Это связано с тем, что нейроны, сообщаются между собой химическими синапсами, в которых есть нейромедиаторы (именно они передают сигнал), но запасы их невелики и могут истощаться, а синтезируются они долго, поэтому при длительной и интенсивной работе нервного центра и, соответственно, нейронов запасы нейромедиатора в синапсе истощаются и тот почти прекращает передачу сигнала – наступает утомление.
Высокая чувствительность к гипоксии. Чем она обусловлена? Нервная ткань потребляет огромное количество энергии, но не имеет запасающих источников этой энергии типа гликогена или ТАГ (нейтральный жир), поэтому вынуждена постоянно получать энергию, окисляя глюкозу крови, хотя ещё можно лактат и кетоновые тела, но в основном глюкозу, причём на процессы этого окисления требуется много кислорода. В случае гипоксии кислорода не хватает, поэтому нервная ткань не может окислять достаточное количество глюкозы, поэтому нервная ткань не получает достаточное количество энергии, а запасающих энергию веществ у неё почти нет, поэтому в нервной ткани наступает гипоэнергетическое состояние. Если обычные ткани могут прекратить свою работу на время нехватки энергии (вот если вы бегать устали, вы же можете просто остановиться), а нервная ткань так не может, ибо без неё организм не управляется, поэтому она работает даже в условиях гипоэнергетики и гипоксии, что, правда, приводит в дальнейшем к нарушению функций и некрозу. Короче говоря, нервной ткани нужно постоянно много энергии, которую она не может получать без кислорода, поэтому она так чувствительна к гипоксии.
Высокая чувствительность к ацидозу и алкалозу. Подробно вопросы алкалоза и ацидоза разбираются в курсе биохимии (биохимия крови и не только) и в курсе нормальной физиологии в разделе физиологии крови, но сейчас вам нужно знать только то, что ацидоз снижает сродство гемоглобина к кислороду, а алкалоз повышает сродство гемоглобина к кислороду. При нормальном сродстве гемоглобин полностью насыщается кислородом в лёгких, а в тканях полностью его отдаёт. Если сродство снижено, то гемоглобин в лёгких будет насыщаться кислородом не полностью, следовательно, и до тканей он будет доносить меньше кислорода. Если сродство выросло, то гемоглобин в тканях отдаёт не весь кислород (сродство высокое – гемоглобин не хочет отдавать кислород). В обоих случаях тканям не хватает кислорода, а это по сути гипоксия, а чем опасна гипоксия для нервных центров, мы уже сказали.
Высокая чувствительность к ядам. Здесь мы снова возвращаемся к теме синапсов. Дело в том, что ряд ядов либо блокирует, либо активирует рецепторы нейромедиаторов на постсинаптической мембране, нарушая передачу нервного сигнала. Например, столбнячный токсин блокирует все рецепторы тормозных нейромедиаторов – в нервной системе вообще нет торможения, поэтому любое возбуждение распространяется, куда ему вздумается, никакой координации нет – всё возбуждено, поэтому у человека максимально напряжены все мышцы, но больше разгибатели, поэтому человек встаёт в позу мостика, но мышцы так сильно сжимаются, что ломают кости, плюс любое прикосновение к человеку сулит ему сильной болью. Есть и другие яды: стрихнин блокирует глициновые рецепторы, яд ботулизма блокирует все рецепторы возбуждающих нейромедиаторов.
Пластичность. Очень интересное и жизненно необходимое свойство нервных центров. Это способность нейронов разрушать старые и образовывать новые связи, перестраиваясь с целью обучения и адаптации. Помимо этого, пластичность может выражаться в отмирании нейронов, что делается для освобождения места для проложения новых путей сообщения между нейронами (отростков).
Закономерности проведения возбуждения по рефлекторной дуге.
Вот мы уже обсудили множество структур в ЦНС, их свойства, особенности, а теперь нужно всё обобщить и поговорить о закономерностях проведения возбуждения по рефлекторной дуге, их 6: одностороннее проведение, замедленное проведение, суммация подпороговых раздражителей, трансформация ритма возбуждения, рефлекторное последействие, посттетаническая потенциация. Разберёмся в каждом отдельно.
Одностороннее проведение. Помните, что если раздражать периферический нерв, то возбуждение пойдёт в обе стороны? Так вот в ЦНС всё несколько иначе: возбуждение возникает в аксональном холмике и идёт от него по аксону к его терминалиям и синапсам. Но почему возбуждение не распространяется на тело, дендриты и не передаётся через синапсы на другие нейроны, от которых обычно приходит сигнал? А дело в том, что в химических синапсах сигнал передаётся только в одну сторону. Сигнал передаётся нейромедиаторами, они же выделяются только из пресинаптической мембраны, а рецепторы к ним есть только в постсинаптической мембране, поэтому сигнал может идти только в одну сторону (постсинаптическая мембрана не выделяет нейромедиаторы, а пресинаптическая мембрана их не воспринимает).
Замедленное проведение. Сами нейроны проводят сигнал (возбуждение) очень быстро, но соединяются между собой они химическими синапсами, которые проводят сигнал с очень большой (по меркам скорости распространения возбуждения по нервным волокнам) задержкой, поэтому прохождение сигнала по рефлекторной дуге занимает время из-за этих задержек.
Суммация подпороговых раздражителей. Дело в том, что нейрон обычно не принимает на себя ПД от других нейронов, нет, другие нейроны в синапсах запускают образование в нейроне ВПСП и ТПСП. Выходит, что другие нейроны по отношению к нашему выступают в роли раздражителей, причём подпороговых, так как ПД они не вызывают. Зато наш нейрон способен суммировать эти раздражения. Рассмотрим суммацию ВПСП. Она бывает последовательной и одновременной. При последовательной на нейрон поступает несколько раздражений с определённой частотой. Если частота высокая, то промежутки между раздражениями небольшие, раздражения вызывают локальные ответы, т.е. деполяризации медленные, которые накладываются друг на друга и достигают Е-критического, вызывая ПД. При одновременной суммации несколько раздражений происходят одновременно, поэтому они действуют вместе, вызывая полноценную медленную деполяризацию, достигающую Е-критического и позволяющую генерировать ПД. На рисунке слева последовательная (раздражения в течение времени, вслед друг за другом), справа одновременная (несколько раздражений в один момент времени) суммации.

Трансформация ритма возбуждения. Под ритмом будем понимать частоту и количество возбуждений. Мы можем уменьшать и увеличивать ритм. Уменьшаем ритм за счёт существования синапсов, они существенно снижают лабильность и частоту из-за синаптической задержки. А вот повышать ритм мы можем за счёт длительной следовой деполяризации, полисинаптических и циркуляционных цепей.

На графике ПД нейрона я изобразил длительную следовую деполяризацию (следовой гиперполяризации нет, но вы не удивляйтесь – она далеко не у всех клеток есть), ей соответствует фаза вторичной экзальтации, т.е. повышенной возбудимости, поэтому нейрон может возбуждаться подпороговыми раздражителями, которыми обычно не раздражается, это приведёт к внеплановому появлению новых ПД, т.е. число импульсов увеличивается за счёт повышения возбудимости, которое обусловленно спецификой калиевых каналов данных нейронов. Что касается полисинаптических сетей, то это сочетание дивергентной и конвергентной нейронных сетей. Суть в том, что на входной нейрон поступает 1 сигнал, который разделяется на 3 таких же, которые отходят на другие нейроны, но затем эти 3 сигнала возвращаются на выходной нейрон, но не одновременно, а последовательно (нейроны с разной скоростью передавали сигнал), поэтому выходной нейрон генерирует 3 сигнала, повышая количество и частоту сигналов. Что касается циклической цепи, то её мы уже обсуждали – из-за циркуляции сигнала один сигнал на входе даёт множество сигналов на выходе – ритм увеличен.
Рефлекторное последействие. Начнём с примера. Раздражитель действует 2 секунды, ответная реакция наступает спустя 1 секунду и длится 2 секунды, весь опыт длится 3 секунды. В течение первой секунды есть раздражение рецептора в рефлекторной дуге, а ответа нет – синаптические задержки (из-за них ответ долго ждать), в течение второй секунды есть и раздражение, и ответ (логично всё), в течение третьей секунды раздражения уже нет, а ответ продолжается – вот то, что происходит в третью секунду, и называется рефлекторным последействием – продолжение ответной реакции в отсутствие раздражителя. Рефлекторное последействие, как и положительная трансформация ритма возбуждения, обусловлена длительной следовой деполяризацией нейронов (основной раздражитель ушёл, но подпороговые раздражители продолжают порождать ПД, нервные импульсы и ответную реакцию, пользуясь повышенной возбудимостью), полисинаптическими сетями (вы на них один раз раздражающе подействовали и перестали, а потом они вам выдают 5 ПД и длительный ответ на основе этих ПД), циклическими сетями (ну тут мы уже говорили, одно раздражение, а сеть порождает сотни и даже больше ПД какое-то время, т.е. раздражать перестали, а ПД и ответные реакции всё идут и идут).
Посттетаническая потенциация. Великая вещь, играет большую роль в научении и запоминании. Если вы вспомните принцип работы химического синапса, то экскрецию нейромедиатора там запускалась входом в клетку кальция, который затем выкачивался из клетки. Но если вы часто используете конкретный синапс, то кальций входит в клетку часто и выкачиваться из неё не успевает, т.е. когда на аксон придёт очередное возбуждение, то в клетку надо будет закачать уже не так много кальция, потому что оттуда ещё прежний выкачать не успели! Плюс ещё митохондрии копят кальций. Выходит, что если вы часто используете какой-то синапс, то проведение в нём сигнала облегчается и ускоряется (синаптическая задержка снижается, так как меньше времени нужно на закачку в клетку кальция), тогда сигнал между конкретными нейронами пробегает значительно быстрее, чем между другими, поэтому в будущем для конкретной ситуации будет использоваться именно эта цепочка нейронов, как более успешная (по ней сигнал легче проходит). Как это работает? Разберём на абстрактном примере. Вы решаете задачу, информация о которой поступает на вход в систему, блок управления формирует три способа решения, каждый способ решения – это отдельная нейронная цепочка/сеть. Сигнал по каждой из трёх цепей проходит с примерно одинаковой скоростью. Но из трёх решений верно только одно, система пробует применить каждое из решений, обратная связь говорит системе, что эффективным оказалось только второе решение, система это понимает и в дальнейшем вновь использует второе решение, т.е. посылает сигнал по второй нейронной цепочке, и делает это ни раз, в её синапсах формируется посттетаническая потенциация, скорость проведения по этой цепочке увеличивается. Спустя какое-то время система вновь сталкивается с этой задачей, посылает небольшой импульс на все три цепочки, но на второй цепочке скорость гораздо выше, поэтому она первой выдаёт решение, его система и использует. Посттетаническая потенциация формирует память, опыт, обучение именно по такому принципу. Вот поэтому, если вы повторяете материал во время его изучения, он хорошо запоминается, но если повторять слишком часто, то он забудется. Почему? Потому что при слишком частом использовании синапсов в них истощаются запасы нейромедиаторов и наступает утомление, поэтому надо знать середину. Кратковременный эффект посттетанической потенциации заключается в том, что кальций не успевает выходить из клетки при частом использовании, а долговременный заключается в том, что кальций накапливается в митохондриях и в случае чего они готовы его вернуть в цитоплазму.
Торможение в ЦНС.
До этого момента мы обсуждали возбуждение в ЦНС, а теперь настало время обсудить такой важнейший и интересный процесс, как торможение. Определение этого понятия вы должны знать очень хорошо! Торможение – самостоятельный, активный, обратимый процесс подавления возбуждения, вызванный возбуждением. Разберём определение по составляющим. Процесс является активным, потому что предполагает затраты энергии на своё осуществление. Процесс самостоятельный в том смысле, что он не является простым следствием другого процесса, а сам имеет определённое значение и выполняет определённые функции, не является вторичным по отношению к чему-либо. Он обратим, т.е. его действие всегда временно (ибо то, что он тормозит, нам ещё ни раз понадобится, поэтому нет смысла это затормаживать раз и навсегда). Торможение названо так, потому что тормозит/подавляет возбуждение в одном месте, но само торможение включается другим возбуждением.
Пару слов об истории открытия возбуждения. Открыл торможение Сеченов. Он сделал таламический препарат лягушки, подвесил лягушку, опустил её лапку в кислоту, через 3 секунды лапка отдёргнулась – это защитный рефлекс, а рефлекс – это возбуждение, идущее по рефлекторной дуге. Затем он положил на таламус кристалл поваренной соли, снова опустил лапку в кислоту, лягушка отдёрнула лапку спустя 15 секунд – на лицо замедление/торможение рефлекса, а значит, и торможение/подавление возбуждения. Потом Сеченов убрал кристалл соли и спустя некоторое время повторил эксперимент – снова 3 секунды, как и до кристалла – обратимость. Таким образом, Сеченов узнал, что торможение – это обратимый процесс подавления возбуждения. Затем Гольц провёл эксперимент с спинальным препаратом лягушки. Что он делал? Тоже опускал лапку в кислоту – лягушка отдёргивала лапку за 3 секунды - защитный рефлекс, а потом он очень сильно зажимал вторую лапку, серьёзно раздражая в ней болевые рецепторы, порождая другой рефлекс (раздражение рецепторов пороговым и сверхпороговым раздражителями предполагает в дальнейшем формирование рефлекса), затем он снова засовывал свободную лапку в кислоту, но та отдёргивалась спустя 15 секунд – снова торможение. Но что нового в этом эксперименте? А то, что один рефлекс подавлял другой, а рефлексы – это возбуждение, идущее по рефлекторным дугам. Но здесь ведь имело место торможение, получается, что одно возбуждение тормозит другое, т.е. торможение одного возбуждения активируется другим возбуждением. Понимание того, что торможение – это активный и самостоятельный процесс, пришло позже. К слову, Сеченов, когда открыл торможение, даже не подозревал, насколько важно его открытие. О значении торможения мы поговорим чуть позже, но сейчас просто скажу, что вся нервная ЦНС работает на сочетании возбуждения и торможения, т.е. Сеченов открыл целый пласт в физиологии!
Разберёмся в механизмах торможения. Выделяют первичные и вторичные. Первичные виды торможения осуществляются специальными тормозными нейронами. Вторичное торможение обусловлено свойствами нейрона.
К первичному виду торможения относят два механизма: пресинаптическое и постсинаптическое торможение.

Пресинаптическое торможение. Осуществляется необычным образом, так как тут имеется два синапса – тормозящий и возбуждающий. Тормозной синапс является аксо-аксональным (между красным и синим). Тормозной нейрон вызывает длительную деполяризацию мембраны «синего» нейрона, в результате такой длительной деполяризации часть натриевых каналов становится инактивированными, их нельзя включить, если на «синий» нейрон придёт волна возбуждения, то этот участок мембраны не возбудится, ибо часть его натриевых каналов не будет работать вследствие инактивации натриевых каналов длительной деполяризацией. Торможение называется пресинаптическим, потому что торможение осуществляется за счёт снижения возбудимости пресинаптической мембраны возбуждающего синапса. Постсинаптическое торможение. Здесь существует только тормозной синапс, он работает, как и все химические синапсы, но тормозной нейрон выделяет тормозной нейромедиатор, который в постсинаптической мембране открывает каналы для калия и хлора. Хлор входит (входят «минусы»), калий выходит (уходят «плюсы») – потенциал возрастает в отрицательную сторону, образуется истинная гиперполяризация (ТПСП), которая снижает возбудимость (расстояние до Е-критического выше, пороговый потенциал выше, возбудить мембрану сложнее – понижается возбудимость) постсинаптической мембраны, поэтому это торможение и называют постсинаптическим. Этот механизм торможение называют «истинным», ибо он является основным, значительно преобладая над другими видами.
Теперь о вторичных механизмах торможения, к ним относятся торможение вслед за возбуждением (постактивационное торможение) и пессимальное.
Постактивационное заключается в том, что у нейрона в ПД очень длинная следовая гиперполяризация (в неё возбудимсть снижается серьёзно, ибо расстояние до Е-критического увеличивается и пороговый потенциал растёт), т.е. нейрон возбудился, а потом затормозился.

Пессимальное торможение срабатывает при высокой частоте раздражения, стимулы попадают на мембрану, когда та находится в фазе относительной рефрактерности (сниженная возбудимость), поэтому каждый новый ПД теряет в амплитуде, пока ПД просто не прекратятся.
Помимо механизма, торможение классифицируют по архитектуре нейронных сетей с участием тормозных нейронов (речь о постсинаптическом торможении), выделяют циркулярное, рецепрокное и латеральное торможение (ну это основные виды).
Циркулярное торможение. Мы уже ни раз говорили о циркулярных нейронных сетях, тормозный нейрон в них ограничивает работу цикла, без него бы возбуждение циркулировало без конца, что нарушало бы адекватную работу ЦНС.

Рецепрокное торможение. Я привёл две допустимые схемы. Суть состоит в том, что есть две рефлекторные дуги, в каждой условно выделим ключевой нейрон, ключевые нейроны тормозят друг друга через тормозные нейроны взаимно. Что это даёт? Такая схема позволяет сделать так, что если возбуждается один, то тормозится другой и наоборот, но никогда они не будут возбуждены вместе одновременно. Принцип рецепрокности помогает регулировать антагонистические процессы, такие как дыхание (регулирую цикл вдох-выдох, нейроны дыхательного центра по очереди друг друга тормозят) и ходьба (пока одна нога стоит, вторая делает шаг, потом наоборот).

Латеральное торможение. Интересный вид торможения – возбудившийся нейрон тормозит через тормозные нейроны себе подобные другие нейроны.

Это можно встретить в сетчатке глаза. Это позволяет сделать картинку чёткой/резкой. Как? Ну вот объект расположен над одним нейроном непосредственно, нейрон генерирует интенсивные сигналы, над вторым нейроном (соседним) предмет расположен как бы споку и действует на него не так сильно, поэтому второй нейрон посылает менее интенсивные сигналы, а третий ещё менее интенсивные. Получается, что предмет на его границе выходит размазанным (ибо мозг видит небольшую разницу между данными первого, второго и третьего нейрона) и не имеет чёткие контуры, а вот если первый нейрон затормозил через тормозной нейрон первый нейрон, то импульсации от второго не будет и информации о размазанной границе не будет – будет информация только с первого нейрона (высокая интенсивность) и от третьего (крайне низкая интенсивность), между ними наш мозг увидит чёткую разницу, поэтому и на картинке границы объекта будут чёткими и неразмытыми.
Чем важно для нас торможение? Каковы его функции? Во-первых, оно координирует рефлекторные акты. Что это значит? У рефлекса может быть несколько вариантов развития, а какой именно вариант будет, решают тормозные нейрон, тормозя неподходящие в конкретных условиях пути и не тормозя подходящие пути. Во-вторых, торможение ограничивает иррадиацию. Иррадиация – распространение возбуждения по ЦНС на множество нервных центров. Дело в том, что возбуждающие нейроны имеют между собой очень обширные связи, поэтому сигнал со всего одного нейрона может распространиться на немалую часть ЦНС, но это далеко не всегда нам нужно, поэтому надо как-то регулировать распространение возбуждения – это делается посредством его торможения. В-третьих, торможение выполняет защитную функцию, предохраняя нейроны от перевозбуждения.
Координация в ЦНС.
Теперь поговорим об очень важном понятии, с которым мы не раз сталкивались – координация. Координация – согласованная последовательная деятельность всех нервных центров при правильной работе ЦНС. А иначе говоря, координация – это правильное взаимодействие и взаимоотношение возбуждения и торможения в ЦНС. Существует 8 принципов координационной деятельности: рецепрокность, иррадиация, общий конечный путь, принцип доминанты, принцип субординации, принцип обратной связи, принципы облегчения и окклюзии.
О рецепрокности мы уже говорили – данный принцип координирует работу антагонистов, обеспечивая поочерёдную их работу, не давая им включаться одновременно. В основе лежит рецепрокная нейронная сеть с участием тормозных нейронов.
Иррадиация – распространение возбуждения по ЦНС, т.е. от одного рецептора, например, импульс может пойти в несколько нервных центров, так как эта информация принципиально важна для нескольких центров, в основе лежит дивергентная нейронная сеть.
Общий конечный путь. Деятельность многих эффекторов контролируется несколькими нервными центрами, а к эффектору ведёт один путь, поэтому приходится создавать конвергентную нейронную сеть, которая собирает информацию с нескольких центров/нейронов, объединяет её (синтез), принимает решение и отправляет сигнал на эффектор.
Принцип доминанты. Доминанта – временно господствующая функциональная система с повышенной возбудимостью. Свойства: повышенная возбудимость, стойкое и инертное возбуждение, способность суммировать возбуждения и тормозить побочные. Грубо говоря, доминанта позволяет нам сконцентрироваться на конкретной задаче и приложить все имеющиеся средства к её решению, дабы полезный результат был достигнут с наиболее высокой вероятностью. Т.е. мы не распыляемся, а решаем проблемы по одной. Принцип доминанты используют и в психологии для воздействия на поведение человека. Приведу утрированный, но понятный пример. Человек умеет плавать, но боится это делать (фобия), фобии в поведении имеют характер доминанты долгое время, поэтому от них сложно избавиться. Что мы сделаем? Посадим человека в лодку, поплывём по озеру (на лодке ему не так страшно, как в воде), затем мы подожжём лодку, а человеку придётся прыгать в воду, ибо доминантой станет желание выжить, а доминанта подавляет все остальные побочные возбуждения, т.е. тормозит рефлексы, в том числе фобию – человек прыгает и плывёт. Одну доминанту мы вытеснили другой, после этого фобия «пошатнулась» и человеку становится легче делать шаги к избавлению от неё, так как фобию наконец поставили под сомнение, сдвинув её с доминантной роли, поэтому она теряет своё «влияние».
Принцип субординации означает, что ЦНС это не простое собрание нервных центров, нет, между этими центрами есть иерархия, позволяющая рационально «распределять обязанности» и осуществлять контроль за работой одних центров над другими.
Принцип обратной связи предполагает существование дополнительных рецепторов, отчитывающихся перед нервными центрами о достигнутом результате, а нервные центры адаптируются и учатся, перестраиваясь в случае, если после их команды не был достигнут положительный результат, опираются они на отчёт от обратной связи, которая может быть положительной или отрицательной.
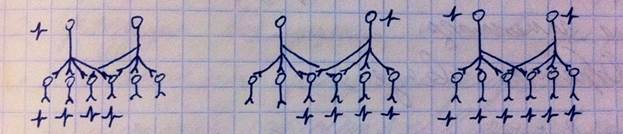
Облегчение. Это совмещённые дивергентные сети. Смотрите на рисунок: есть верхние и нижние нейроны, каждый верхний иннервирует 4 нижних, у каждого верхнего есть 2 собственных и 2 общих. Если возбуждается только один из верхних, он возбуждает только собственные 2 нейрона, а общие не может, ибо у тех низкая возбудимость, зато если включаются оба верхних нейрона, то они активируют все 6 (их совместного эффекта хватает, чтобы возбудить общие нейроны). Т.е. облегчение – это когда сумма эффектов двух нейронов, если бы они работали по отдельности (2 + 2 = 4), меньше, чем результат их совместной работы (6 возбуждённых нижних нейронов).

Окклюзия. Схема та же, но на этот раз общие нижние нейроны могут возбуждаться и одним верхним, и двумя. Т.е. сумма эффектов верхних нейронов нейронов, если бы они работали по отдельности (4 + 4 = 8), больше, чем их совместный эффект (равен 6 активированным нижним нейронам).
Отделы ЦНС.
До этого мы говорили об общих принципах в ЦНС, а теперь необходимо немного обсудить частную физиологию ЦНС, т.е. за что отвечают её конкретные отделы, какое существует функциональное. Конечно, по этой теме можно спокойно писать отдельную книгу, настолько тут много информации, но с этим вы познакомитесь на кафедре неврологии или самостоятельно, наша же задача на курсе нормальной физиологии сформировать общее представление по частной физиологии ЦНС. Начнём с отделов ЦНС.
Спинной мозг необходим для проведения информации с рецепторов в вышележащие отделы ЦНС, информация с некоторых рецепторов минует спинной мозг, ибо передаётся не на спиномозговые нервы, а на черепные (идут сразу в головной мозг), помимо этого, мозг проводит сигналы от вышележащих отделов ЦНС к органам-эффекторам, т.е. две эти функции можно объединить и назвать связующей функцией спинного мозга (связует головной мозг и органы с тканями), но при этом спинной мозг способен осуществлять регуляцию некоторых рефлекторных актов, ибо существуют рефлекторные дуги, замыкающиеся на уровне спинного мозга (пример: коленный рефлекс и ахилов рефлекс, которые кстати являются клинически значимыми в неврологии), ещё мы можем сказать, что спинной мозг обеспечивает спинальный тонус мышц (об этом подробнее в теме мышечного тонуса).
Продолговатый мозг. Структура простая, но очень важная. Из всех отделов головного мозга продолговатый является самым древним, а потому в нём сосредоточена регуляция наиболее важных физиологических процессов – в продолговатом мозге есть нервные центры, регулирующие деятельность сердца, кровеносных сосудов, желудочно-кишечного тракта, также там расположен дыхательный центр. Помимо этого, конечно, через продолговатый мозг проходит множество проводящих путей.
Мост. Участвует в восприятии звука и света, регулирует мимическую мускулатуру и жевательные мышцы, помимо этого через него проходит очень большое количество проводящих путей, мостом он не просто так назван.
Средний мозг. Его основные функции заключаются в осуществлении реакций, связанных со зрением (содружественное движение глазных яблок), обеспечивает реакции тела на свет и звук, а также принимает большое участие в регуляции мышечного тонуса (чёрная субстанция и красное ядро, о них подробнее в теме мышечного тонуса).
Мозжечок. Регулирует и поддерживает мышечный тонус, координирует позные и целенаправленные движения (например, мы можем одновременно шагать и поддерживать вертикальное положение тела, без мозжечка два этих процесса совмещались бы очень плохо), значительно увеличивает точность быстрых целенаправленных движений.
Таламус. В таламус попадает вся информация с органов чувств, его задача заключается в обработке этой информации и отправке её в соответствующие отделы коры больших полушарий (далеко не вся информация от органов чувств попадает в кору, много лишней информации отсортировывается, иначе бы кора была перегружена и не могла бы адекватно работать), помимо этого, таламус участвует в регуляции моторики скелетных мышц (сигнал идёт от коры, но через таламус), ещё таламус поддерживает внимание, память, сознание, является высшим центром боли и надсегментным центром некоторых рефлексов.
Гипоталамус. Чрезвычайно важная и многофункциональная структура! Он является высшим центром вегетативной нервной системы (это вся непроизвольная нервная деятельность), т.е. гипоталамус обеспечивает регуляцию организма с целью поддержания гомеостаза, помимо этого, гипоталамус формирует эмоциональное и мотивационное поведение, вместе с эпифизом регулирует циркадный ритм, через гипофиз осуществляет контроль нервной системой над эндокринной, также гипоталамус осуществляет интеграцию соматической и вегетативной нервной системы (позволяет влиять коре на деятельность внутренних органов, например, при сильном волнении у нас может повыситься ЧСС и АД, но ещё возможно влияние и в обратную сторону, это как раз то самое формирование мотивационного и эмоционального поведения).
Виды нервных систем и их особенности.
Функционально нервную систему можно разделить на соматическую и вегетативную. Чем они отличаются?
| Параметры | Соматическая НС | Вегетативная НС |
| Уровень контроля | Произвольная (подвластна сознанию) | Непроизвольна (мало подвластна) |
| Область иннервации | Органы чувств и скелетная мускулатура | Практически все органы и ткани |
| Строение | Сегментарное | Очаговое |
| Рефлекторные дуги | Рецептор, афферентный, вставочный (ноль/один/много), эфферентный нейроны, эффектор | Рецептор, афферентный, преганглионарный и постганглионарный нейроны, эффектор |
| Рецепторы | Органы чувств, экзорецепторы, пропреорецепторы | Экзо- и эндорецепторы |
Немного пояснений к таблице. Мы говорим, что вегетативная нервная система является непроизвольной, но немного подчиняется нашему сознательному контролю по той причине, что гипоталамус посылает некоторые сигналы от коры больших полушарий на нижележащие нервные центры в составе вегетативной нервной системы (когда будем разбирать регуляцию конкретных физиологических функций, вы это не раз увидите). Что значит сегментарное и очаговое строение? Да в принципе то и значит, что соматическая НС имеет сегменты (повторяющиеся участки с похожим строением), причём все сегменты соединены (не только проводящими путями, но и сами непосредственно), а вегетативная НС разбросана очагами по всей НС. С соматической и вегетативной рефлекторными дугами вы уже знакомились ранее. Что касается рецепторов, экзорецепторы воспринимают информацию из внешней среды (расположены преимущественно на коже, слизистых), эндорецепторы воспринимают информацию из внутренней среды (обычно речь про кровь), проприорецепторы воспринимают информацию из опорно-двигательной системы (мышцы и сухожилия). Соматическая НС принимает информацию от органов чувств и экзорецепторов, так как отвечает за связь организма с внешней средой, также она принимает информацию с проприорецепторов, ибо отвечает за движение тела в пространстве. Вегетативная НС воспринимает информацию с эндорецепторов, так как она должна поддерживать гомеостаз (постоянство внутренней среды), но ещё она воспринимает информацию с некоторых экзорецепторов, ибо гомеостаз необходимо выстраивать в адекватном соответствии внешней среде (например, на улице холодно, чтобы вы не замёрзли, вегетативная НС повысит термогенез и понизит теплоотдачу, но для этого она должна узнать, что на улице холодно – тут-то и понадобятся экзорецепторы, а именно, холодовые терморецепторы кожи).
Вегетативная НС, в свою очередь, функционально делится на симпатическую и парасимпатическую. Хотя дополнительно вполне справедливо выделяют метасимпатическую НС. Чем она отличается? Симпатическая и парасимпатическая НС обязательно проходят через ЦНС, а метасимпатическая не входит в состав ЦНС, по сути метасимпатическая НС представлена рефлекторными дугами, которые замыкаются на уровне нервного ганглия (узла), расположенного около органа или внутри него. Примером таких узлов являются межмышечные нервные сплетения в полых органах желудочно-кишечного тракта, например, кишечник. Благодаря межмышечным сплетениям кишечник способен осуществлять множество движений, необходимых для продвижения и перемешивания пищи в ходе переваривания, т.е. осуществлять перистальтику. Это довольно важная функция, которая может осуществляться без участия ЦНС. Но куда более важными и разнообразными по эффектам является симпатическая и парасимпатическая НС, ибо их охват и функционал гораздо шире, их мы тоже сравним посредством таблицы.
| Параметр | Симпатическая НС | Парасимпатическая НС |
| Время наибольшей активности | Активна во время интенсивной деятельности организма | Более активна во время расслабления организма |
| Основной нейромедиатор | Норадреналин и ему подобные | Ацетилхолин и ему подобные |
| Область иннервации | Внутренние органы, сосуды, железы, кожа, даже мышцы, в общем почти всё | Внутренние органы и некоторые сосуды |
| Строение | Грудной и поясничный отдел спинного мозга | Отделы выше и ниже симпатических |
Некоторые пояснения к таблице. Конечно, симпатическая и парасимпатическая НС в основном являются антагонистами и наибольшую активность приобретают при разных состояниях организма, но обычно они работают совместно (нет такого, чтобы одна из систем полностью отключилась) и они могут являться синергистами иногда.
Двигательная система и её регуляция.
Теперь обсудим систему, контролирующую нашу ориентацию в пространстве и двигательную активность.
Мышечный тонус – это длительное мышечное сокращение без признаков утомления, возникающее под действием ЦНС. Все три компонента этого определения важно запомнить. Зачем нам нужен мышечный тонус? Он позволяет поддерживать и восстанавливать позу. Поза – это определённая относительно постоянная ориентация головы и тела в пространстве.
Какие мышцы участвуют в поддержании тонуса? Для этого давайте обратимся к уже пройденной нами теме частной физиологии мышц. Мышцы прежде всего состоят из мышечных волокон, которые бывают экстрафузальные (сократительные) и интрафузальные (вспомогательные клетки для рецепторов). Экстрафузальные делятся на быстрые/белые и медленные/красные. Так вот красные волокна обладают большими запасами миоглобина и гликогена, поэтому способны сокращаться довольно продолжительно. Мышцы классифицируют в соответствие волокнам, из которых они состоят, если в мышцах преобладают красные волокна, то мышца считается тонической (ну или сильной, медленной, но в этой теме нам больше подходит это слово). Тонус именно этих мышц, а конкретно разгибателей ног, спины и шеи поддерживают нашу позу, противостоя действующей на наше тело гравитации, которая хочет, чтобы мы легли, поэтому тонические мышцы (конкретно эти) иногда называют антигравитационными.
В основе поддержания мышечного тонуса и позы, соответственно, лежат два рефлекса – рефлексы растяжения и сухожильный рефлекс.

Итак, начнём с рефлекса растяжения. На рисунке выше я изобразил мышцу, на ней имеется интрафузальное волокно, которое маленькими «сухожилиями» присоединяется к самой мышце и сухожилию. Если мышца расслабляется и растягивается, то её длина увеличивается, а значит, увеличится и длина интрафузального волокна, а если растёт его длина, то диаметр его уменьшается (логично, ведь объём клетки постоянен). Вокруг интрафузального волокна обмотан дендрит чувствительного нейрона, этот участок дендрита и является рецептором (афферентная часть дуги – чувствительный нейрон с рецептором). Чувствительный нейрон посылает импуьсы на мотонейрон-альфа, который иннервирует данную мышцу (точнее не прям целую мышцу, а несколько волокон, относящихся к нему, эфферентная часть дуги – мотонейрон). Как же работает этот рефлекс? Если мышца растягивается, растягивается и само интрафузальное волокно, рецептор вокруг него это замечает и генерирует нервный импульс, тот идёт по чувствительному нейрону на мотонейрон, который, в свою очередь, посылает импульс на экстрафузальные волокна (сократительные), заставляя мышцу сократиться, растяжения больше нет (мышца же сократилась), поэтому рецептор доволен, рефлекс останавливается (ибо рецептор не раздражается). Так рефлекс контролирует длину мышцы. Как это представить на практике? Вот вы сидите на стуле, немного сутулитесь, под действием гравитации ваше туловище наклоняется вперёд, разгибатели спины при этом пассивно растягиваются (нагнувшееся туловище их растянуло), растяжение было замечено рецепторами, они включили рефлекс, разгибатели сократились и выпрямили вашу спину, рефлекс прекратился, затем вы снова начали наклоняться, потом снова выпрямляться, и так постоянно. Но мы ведь поддерживаем позу, не двигаясь, т.е. не видно, как мы наклоняемся и разгибаемся. На самом деле мы действительно наклоняемся и разгибаемся/выпрямляемся, просто нейроны все тонко настроены и скорость проведения сигнала в них очень большая, поэтому рефлексы эти срабатывают чуть ли не мгновенно, а рецепторы довольно чувствительные, поэтому наши наклоны и разгибания/выпрямления происходят очень быстро и имеют маленькую амплитуду, вот от этого-то мы их и не замечаем (просто мы нагибаемся на 2 мм и через долю секунды выпрямляемся – очень сложно такое заметить). Однако это не всё. Ещё существует мотонейрон-гамма, он получает импульсы от супраспинальных центров (отделы ЦНС над спинным мозгом) и изменяет работу рефлекса растяжения. Как? Мотонейрон-гамма иннервирует интрафузальное волокно, заставляя его сокращаться и изменять тем самым длину и диаметр. Для чего это нужно? Рефлекс растяжения, предохраняя мышцу от растяжения, поддерживает в ней постоянную длину, а постоянная длина в мышцах означает постоянное положение тела в пространстве, т.е. позу. Но в разных позах вам нужно разное положение частей тела, поэтому и мышцам надо иметь уже другую длину, т.е. нужно как-то регулировать длину, которую будет поддерживать мышца. Этим занимается мотонейрон-гамма, который изменяет длину интрафузального волокна, изменяя параметр рефлекса растяжения, чтобы тот поддерживал уже другую длину мышцы.
Второй рефлекс – сухожильный рефлекс. Сухожилие нерастяжимо. Если вы прикладываете к нему усилие, то оно не меняет длину, но в нём накапливается напряжение до тех пор, пока оно не порвётся. И вот, чтобы оно не порвалось, был создан этот рефлекс. В сухожилии есть небольшой пучок, обмотанный дендритом чувствительного нейрона (рецептор), чувствительный нейрон сухожильного рефлекса (афферентной частью дуги является чувствительный нейрон и его рецептор) посылает импульс на тормозный нейрон, который тормозит мотонейрон-альфа (тормозный и мотонейрон-альфа представляют эфферентную часть дуги), чтобы тот не возбуждал мышцу и не заставлял её сокращаться. Как работает рефлекс? Допустим, мышца совершает очень тяжёлую работу, такую, что она грозит разрывом сухожилия – в нём накопилось напряжение, это фиксирует рецептор, он создаёт импульс, импульс идёт по чувствительному нейрону, попадает на тормозный нейрон, тот тормозит мотонейрон-альфа, тот перестаёт возбуждать мышцу, мышца расслабляется, ослабляя напряжение в сухожилии, предохраняя его от разрыва.
Теперь разберёмся в структурах, регулирующих мышечный тонус, на основе которых выделяются соответствующие виды мышечного тонуса: спинальный, бульбарный, мезенцефальный, корковый + подкорковый. Разберёмся во всех по очереди.
Спинальный тонус поддерживается (именно поддерживается, потому что без ЦНС тонус не возможен) спинным мозгом на основе уже знакомых нам рефлекса растяжения и сухожильного рефлекса. Спинальный тонус не позволяет нам поддерживать какую-то стабильную полноценную позу – он лишь создаёт фоновое напряжение мышц. Опыт Броджеста хорошо показывает, что из себя представляет спинальный тонус. В этом опыте используется спинальный препарат лягушки (из ЦНС сохранён только спинной мозг, поэтому из всех тонусов возможен только спинальный), лягушка подвешена на крючок, а её лапки согнуты в коленном суставе, т.е. там есть видимый угол (пускай 135 градусов), затем у одной лапки вы перерезаете седалищный нерв, т.е. тонические мышцы нижней лапки лягушки более не сообщаются со спинным мозгом и спинальный тонус исчезает – угол в коленном суставе увеличивается до 175 градусов. Т.е. спинальный тонус создавал некоторое напряжение, которое в данном случае помогало лапкам противостоять гравитации, но для поддержания полноценной позы этого напряжения было недостаточно, поэтому и говорят, что спинальный тонус создаёт фоновое напряжение.
Бульбарный тонус поддерживается продолговатым мозгом. В нём есть ядро Дейтерса и ретикулярная формация. Ядро Дейтерса возбуждает мотонейроны (в основном гамма, но возможно и альфа) мышц разгибателей в спинном мозге, а те же тормозит мотонейроны мышц сгибателей. Ретикулярная формация, напротив, активирует сгибатели и тормозит разгибатели, выступая как бы в роли антагониста ядра Дейтерса, но влияние ядра Дейтерс сильнее, поэтому в целом продолговатый мозг и, соответственно, бульбарный тонус (обеспечивается спинным и продолговатым мозгом, назван бульбарным, потому что высший отдел – продолговатый мозг) приводят к преобладанию тонуса разгибателей над сгибателями, такой тонус ещё называют контрактильным. Как это будет выглядеть? Физиологи брали кошку (изверги) и из всех отделов головного мозга оставляли только продолговатый, в итоге, у кошки вытягивались лапы и выгибалась (распрямлялась) спина – преобладание тонуса мышц разгибателей.
Мезенцефальный тонус поддерживается средним мозгом. В нём есть красные ядра (если пути от ядра Дейтерса не перекрещиваются, то пути от ядер Дейтерса перекрещиваются), которые активируют мотонейроны сгибателей и тормозят мотонейроны разгибателей, т.е. красное ядро выступает значительным антагонистом ядрам Дейтерса, а средний мозг выступает антагонистом продолговатого, но мезенцефальный тонус предполагает совместное функционирование спинного, продолговатого и среднего мозга, поэтому продолговаты и средний уравнивают друг друга, поэтому бульбарный тонус позволяет поддерживать стабильные нормальные позы. Если кошке оставили средний мозг, то если ей придать какую-либо позу, то она будет её сохранять, поэтому такой тонус ещё называют пластическим.
Теперь давайте обсудим мозжечок. Он не поддерживает конкретный вид тонуса, более того, мозжечок не самый обязательный отдел мозга, в том смысле, что если вы удалите человеку гипоталамус, то он не выживет, а если мозжечок, то он будет жить, но с определёнными трудностями. Каковы функции мозжечка? Во-первых, он регулирует (не поддерживает, а регулирует) мышечный тонус и позы. Во-вторых, он координирует позные и целенаправленные движения (вы ведь не просто поддерживаете позу, вы ведь часто ещё совершаете какие-то целенаправленные движения, например, когда вы ходите, вам нужно одновременно двигаться вперёд и поддерживать вертикальное положение тела, мышцы ног участвуют сразу и в позных, и в целенаправленных движениях, мозжечок делает так, чтобы они нормально согласовались и не мешали друг другу). В-третьих, мозжечок позволяет совершать быстрые целенаправленные движения с высокой точностью. В общем роль мозжечка в мышечном тонусе, да и вообще в работе всей опорно-двигательной системы, хоть он и считается необязательным отделом мозга (если решите какому-нибудь преподавателю сказать, что мозжечок не является обязательным отделом мозга, то не забудьте это аргументировать, а потом подчеркнуть роль мозжечка, ибо он не обязательный, но и не лишний – лишнего в нашем организме ничего не бывает, ну бессмысленного уж точно). Какие заболевания могут развиваться при поражении мозжечка? Атаксия – нарушение координации позных и целенаправленных движений. Дисметрия – утрата размерности (человек не регулирует скорость движения конечностей и расстояние, которое они проходят в процессе движения). Адиадохокинез – нарушение координации деятельности мышц-антагонистов (между ними должно быть равновесие, которое может равномерно смещаться, вместо этого оно смещается неравномерно, поэтому движения становятся отрывистыми и затруднительными). Астения – повышение утомляемости мышц (в следствие неадекватной их работы). Астазия – неспособность мышц к длительному тоническому сокращению и, как следствие, невозможность стоять.
Теперь поговорим о роли коры больших полушарий и базальных ганглиях. Кора не столько поддерживает мышечный тонус, сколько регулирует (нам иногда приходится менять позы и учиться новым, в этом нам помогает кора, ведь она в принципе способна управлять всеми скелетными мышцами. А вот что касается базальных ганглиев (ядер, подкорки)? Торможение чёрной субстанции и медиального бледного шара приводит к тому, что таламус легче возбуждает кору – это облегчает движения, провоцируемые корой. Если же чёрная субстанция и медиальный бледный шар возбуждаются, то таламус слабее действует на кору, в результате ограничиваются ненужные движения.
Настало время рассмотреть установочно-тонические рефлексы, которые являются результатом действия не какого-то конкретного уровня регуляции мышечного тонуса, а результатом действия всей системы поддержания и регуляции мышечного тонуса в целом. Установочно-тонические рефлексы направлены на поддержание определённой позы и равновесия в гравитационном поле Земли. Выделяют статические и статокинетические установочно-тонические рефлексы.
Статические установочно-тонические рефлексы возникают при изменении положения тела, не связанных с перемещением тела в пространстве. Выделяют два типа таких рефлексов: рефлексы позы и выпрямительные рефлексы. Рефлексы позы направлены на сохранение позы при угрозе её потери. Для этого используется информация с пропреорецепторов (рецепторы растяжения и сухожильный орган от мышц и сухожилий шеи) и вестибулорецепторов из вестибулярного аппарата, участвует в основном продолговатый мозг, он перераспределяет работу разгибателей с целью сохранения позы в изменяющихся условиях. Что это значит на практике? Вы стоите, вас толкнули, но вы не упали, а отшагнули и сохранили вертикальное положение тела, сохранив позу при угрозе её нарушения. Выпрямительные рефлексы направлены на восстановление уже утраченной позы. Для этого используется информация с вестибулорецепторов, сетчатки глаз, пропреорецепторы шеи и туловища и даже рецепторы кожи туловища (хотя это уже больше для животных, живущих в норах), эти вопросы уже решает средний мозг. Эти рефлексы последовательно восстанавливают позу тела: сначала голова, потом всё остальное тело. Если вы положите кролика на бок и посмотрите, как он








