 2014-02-05
2014-02-05 1755
1755ГЛАВА 4. РЕПРОДУКЦИЯ ВИРУСОВ
Таксономические признаки вирусов животных
Таблица 7.
| Семейство | Нуклеиновая кислота | Тип симметрии капсида | Наличие оболочки | Размер вириона | ||
| тип | структура | мол. масса | ||||
| Поксвирусы | ДНК | Д | (85-240)·106 | |||
| Герпесвирусы | ДНК | Д | (90-130)·106 | Кубический | + | |
| Гепаднавирусы | ДНК | Д | 1,6·106 | Сложный | + | 45-50 |
| Аденовирусы | ДНК | Д | (20-30)·106 | Кубический | - | 70-90 |
| Паповавирусы | ДНК | О | (3-5)·106 | Кубический | - | 45-55 |
| Парвовирусы | ДНК | ДК | (1,5-2,2)·106 | Кубический | - | 18-26 |
| Реовирусы | РНК | ДФ | (12-19)·106 | Кубический | - | 60-80 |
| Тогавирусы | РНК | О | 4·106 | Кубический | + | 30-90 |
| Коронавирусы | РНК | О | 9·106 | Спиральный | + | 80-130 |
| Парамиксовирусы | РНК | О | 5·106 | Спиральный | + | 150-300 |
| Рабдовирусы | РНК | О | 3·106 | Спиральный | + | 70-175 |
| Ортомиксовирусы | РНК | ОФ | 5·106 | Спиральный | + | 80-120 |
| Буньявирусы | РНК | ОФК | 6·106 | Спиральный | + | 90-100 |
| Аренавирусы | РНК | ОФ | 3·106 | Спиральный | + | 50-300 |
| Ретровирусы | РНК | О | 7·106 | Сложный | + | 80-100 |
| Пикорнавирусы | РНК | О | 2·106 | Кубический | - | 20-30 |
| Калицивирусы | РНК | О | (2,6-2,8)·106 | Кубический | - | 20-30 |
Обозначения: Д — двунитчатая; К — кольцевая; О — однонитчатая; Ф — фрагментированная; «+» — есть; «—» — нет.
Процесс репродукции вирусов может быть условно разделен на две фазы. Первая фаза охватывает события, которые ведут к адсорбции и проникновению вируса в клетку, освобождению его внутреннего компонента и модификации его таким образом, что он способен вызвать инфекцию. Соответственно, первая фаза включает в себя три стадии: 1) адсорбция вируса на клетках; 2) проникновение вируса в клетки; 3) раздевание вируса в клетке. Эти стадии направлены на то, чтобы вирус был доставлен в соответствующие клеточные структуры, и его внутренний компонент был освобожден от защитных оболочек. Как только эта цель достигнута, начинается вторая фаза репродукции, в течение которой происходит экспрессия вирусного генома. Эта фаза включает в себя стадии: 1) транскрипции, 2) трансляции информационных РНК, 3) репликации генома, 4) сборки вирусных компонентов. Заключительной стадией репродукции является выход вируса из клетки.
АДСОРБЦИЯ ВИРУСА НА КЛЕТКАХ
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусных частиц к клеточной поверхности. Процесс адсорбции возможен при наличии соответствующих рецепторов на поверхности клетки и «узнающих»их субстанций на поверхности вируса. Самые начальные процессы адсорбции имеют неспецифический характер, и в основе их может лежать электростатическое взаимодействие положительно и отрицательно заряженных группировок на поверхности вируса и клетки. Однако узнавание клеточных рецепторов вирусными белками, ведущее к прикреплению вирусной частицы к клетке, является высоко специфическим процессом. Белки на поверхности вируса, узнающие специфические группировки на плазматической мембране клетки и обусловливающие прикрепление к ним вирусной частицы, называются прикрепительными белками.
Вирусы используют рецепторы, предназначенные для прохождения в клетку необходимых для ее жизнедеятельности веществ: питательных веществ, гормонов, факторов роста и т. д. Рецепторы могут иметь разную химическую природу и представлять собой белки, углеводный компонент белков и липидов, липиды. Рецепторами для вирусов гриппа и парамиксовирусов является сиаловая кислота в составе гликопротеидов и гликолипидов (ганглиозидов), для рабдовирусов и реовирусов — также углеводный компонент в составе белков и липидов, для пикорнавирусов и аденовирусов — белки, для некоторых вирусов — липиды. Специфические рецепторы играют роль не только в прикреплении вирусной частицы к клеточной поверхности. Они определяют дальнейшую судьбу вирусной частицы, ее внутриклеточный транспорт и доставку в определенные участки цитоплазмы и ядра, где вирус способен инициировать инфекционный процесс. Вирус может прикрепиться и к неспецифическим рецепторам и даже проникнуть в клетку, однако только прикрепление к специфическому рецептору приведет к возникновению инфекции.
Прикрепление вирусной частицы к клеточной поверхности вначале происходит путем образования единичной связи вирусной частицы с рецептором. Однако такое прикрепление непрочно, и вирусная частица может легко оторваться от клеточной поверхности — обратимая адсорбция (рис. 7, а). Для того чтобы наступила необратимая адсорбция, должны появиться множественные связи между вирусной частицей и многими молекулами рецепторов, т. е. должно произойти стабильное мультивалентное прикрепление. Количество молекул клеточных рецепторов в участках адсорбции может доходить до 3000. Стабильное связывание вирусной частицы с клеточной поверхностью в результате мультивалентного прикрепления происходит благодаря возможности свободного перемещения молекул рецепторов в липидном бислое плазматической мембраны, которое определяется подвижностью, «текучестью» белково-липидного слоя. Увеличение текучести липидов является одним из наиболее ранних событий при взаимодействии вируса с клеткой, следствием которого является формирование рецепторных полей в месте контакта вируса с клеточной поверхностью и стабильное прикрепление вирусной частицы к возникшим группировкам — необра
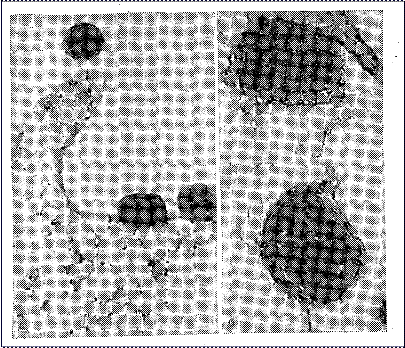 |
тимая адсорбция (рис. 7, б).
Рис. 7. Адсорбция вируса осповакцины на клетке (электронно-микроскопическое изображение): а – обратимая; б – необратимая.
Количество специфических рецепторов на поверхности клетки колеблется между 104 и 105 на одну клетку. Рецепторы ряда вирусов могут быть представлены лишь в ограниченном наборе клеток-хозяев, и этим может определяться чувствительность организма к данному вирусу. Например, пикорнавирусы адсорбируются только на клетках приматов. Рецепторы для других вирусов, напротив, широко представлены на поверхности клеток различных видов, как, например, рецепторы для ортомиксовирусов и парамиксовирусов, представляющие собой сиалил-содержащие соединения. Поэтому эти вирусы имеют относительно широкий диапазон клеток, на которых может происходить адсорбция вирусных частиц. Рецепторами для ряда тогавирусов обладают клетки исключительно широкого круга хозяев: эти вирусы могут адсорбироваться и инфицировать клетки как позвоночных, так и беспозвоночных.
Наличие специфических рецепторов на поверхности клетки в ряде случаев обусловливает феномен зависимого от хозяина ограничения, т. е. способность вируса заражать лишь определенные виды животных. В целом ограничения при взаимодействии рецепторных систем вируса и клетки биологически оправданы и целесообразны, хотя в ряде случаев они являются «перестраховкой». Так, многие линии клеток, устойчивых к вирусам полиомиелита и Коксаки, можно заразить депротеинизированными препаратами РНК, выделенными из этих вирусов. Такое заражение клеток идет в обход естественных входных путей инфекции через взаимодействие с клеточными рецепторами. Известна потенциальная способность вирусов животных реплицироваться в протопластах дрожжей, грибов и бактерий, а бактериофагов — в клетках животных. Таким образом, вирусные ДНК и РНК обладают способностью заражать и более широкий круг хозяев, чем вирусы.
ПРОНИКНОВЕНИЕ ВИРУСОВ В КЛЕТКУ
Исторически сложилось представление о двух альтернативных механизмах проникновения в клетку вирусов животных — путем виропексиса (эндоцитоза) и путем слияния вирусной и клеточной мембран. Однако оба эти механизма не исключают, а дополняют друг друга.
Термин «виропексис», означает, что вирусная частица попадает в цитоплазму в результате инвагинации участка плазматической мембраны и образования вакуоли, которая содержит вирусную частицу.
Рецепторный эндоцитоз. Виропексис представляет собой частный случай рецепторного или адсорбционного эндоцитоза. Этот процесс является обычным механизмом, благодаря которому в клетку поступают питательные и регуляторные белки, гормоны, липопротеины и другие вещества из внеклеточной жидкости. Рецепторный эндоцитоз происходит в специализированных участках плазматической мембраны, где имеются специальные ямки, покрытые со стороны цитоплазмы особым белком с большой молекулярной массой — клатрином. На дне ямки располагаются специфические рецепторы. Ямки обеспечивают быструю инвагинацию и образование покрытых клатрином внутриклеточных вакуолей. Полупериод проникновения вещества внутрь клетки по этому механизму не превышает 10 мин с момента адсорбции. Количество образующихся в одну минуту вакуолей достигает более 2000. Таким Образом, рецепторный эндоцитоз представляет собой хорошо слаженный механизм, который обеспечивает быстрое проникновение в клетку чужеродных веществ.
Покрытые вакуоли сливаются с другими, более крупными цитоплазматическими вакуолями, образуя рецептосомы, содержащие рецепторы, но не содержащие клатрин, а те в свою очередь сливаются с лизосомами. Таким путем проникшие в клетку белки обычно транспортируются в лизосомы, где происходит их распад на аминокислоты; они могут и миновать лизосомы, и накапливаться в других участках клетки в недеградированной форме. Альтернативой рецепторного эндоцитоза является жидкостный эндоцитоз, когда инвагинация происходит не в специализированных участках мембраны. Большинство оболочечных и безоболочечных вирусов животных проникает в клетку по механизму рецепторного эндоцитоза. Эндоцитоз обеспечивает внутриклеточный транспорт вирусной частицы в составе эндоцитарной вакуоли, поскольку вакуоль может двигаться в любом направлении и сливаться с клеточными мембранами (включая ядерную мембрану), освобождая вирусную частицу в соответствующих внутриклеточных участках. Таким путем, например, ядерные вирусы попадают в ядро, а реовирусы — в лизосомы. Однако проникшие в клетку вирусные частицы находятся в составе вакуоли и отделены от цитоплазмы ее стенками.Им предстоит пройти ряд этапов, прежде чем они смогут вызвать инфекционный процесс.
Слияние вирусной, и клеточной мембран. Для того чтобы внутренний компонент вируса мог пройти через клеточную мембрану, вирус использует механизм слияния мембран. У оболочечных вирусов слияние обусловлено точечным взаимодействием вирусного белка слияния с липидами клеточной мембраны, в результате которого вирусная липопротеидная оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса оказывается по другую ее сторону. У безоболочечных вирусов один из поверхностных белков также взаимодействует с липидами клеточных мембран, в результате чего внутренний компонент проходит через мембрану. Большинство вирусов животных выходит в цитозол из рецептосомы.
Если при эндоцитозе вирусная частица является пассивным пассажиром, то при слиянии она становится активным участником процесса. Белком слияния является один из ее поверхностных белков. К настоящему времени этот белок идентифицирован лишь у парамиксовирусов и ортомиксовирусов. У парамиксовирусов этот белок (Р-белок) представляет собой один из двух гликопротеидов, находящихся на поверхности вирусной частицы. Функцию белка слияния у вируса гриппа выполняет малая гемагглютинирующая субъединица.
Парамиксовирусы вызывают слияние мембран при нейтральном рН, и внутренний компонент этих вирусов может проникать в клетку непосредственно через плазматическую мембрану. Однако большинство оболочечных и безоболочечных вирусов вызывают слияние мембран только при низком значении рН — от 5,0 до 5,75. Если к клеткам добавить слабые основания (хлорид аммония, хлороквин и др.), которые в эндоцитарных вакуолях повышают рН до 6,0, слияния мембран не происходит, вирусные частицы остаются в вакуолях, и инфекционный процесс не возникает. Строгая зависимость слияния мембран от значений рН обусловлена конформационными изменениями вирусных белков слияния.
В лизосоме постоянно имеется низкое значение рН (4,9). В эндоцитарной вакуоли (рецептосоме) закисление создается за счет АТФ-зависимого «протонового насоса» еще на клеточной поверхности при образовании покрытой вакуоли. Закисление эндоцитарной вакуоли имеет большое значение для проникающих в клетку физиологических лигандов, так как низкое значение рН способствует диссоциации лиганда от рецептора и рециркуляции рецепторов.
Схематическое изображение возможных способов проникновения вирусов в клетку показано на рис. 8.
 | 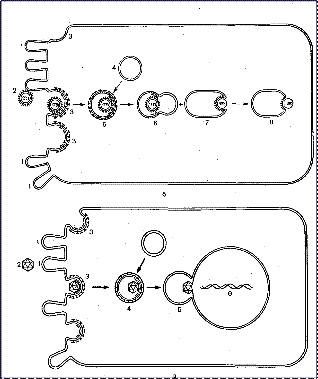 | ||
Рис. 8. Проникновение вируса в клетку:
а — проникновение вируса путем слияния оболочки с плазматической мембраной;
б — проникновение вируса в цитоплазму путем рецепторного эндоцитоза:
1 - клеточная ворсинка;
2 – вирион;
3 - покрытая ямка, содержащая рецепторы;
4 - цитоплазматическая вакуоль;
5 - вакуоль, содержащая вирион;
6 - рецептосома, образовавшаяся в результате слияния покрытой вакуоли с цитоплазматической вакуолью;
7 - слияние вирусной оболочки со стенкой рецептосомы;
8 - выход внутреннего компонента вируса в цитоплазму;
в — проникновение вируса в ядро посредством рецепторного эндоцитоза:
1 - клеточные ворсинки;
2 - внеклеточный вирион;
3 - адсорбция вируса на рецепторах покрытой ямки;
4 - покрытая вакуоль;
5 - образование рецептосомы и ее слияние с наружной ядерной оболочкой;
6 - высвободившаяся вирусная ДНК.
Тот же механизм, который лежит в основе слияния вирусных и клеточных мембран, обусловливает индуцированный вирусами гемолиз и слияние плазматических мембран прилежащих друг к другу клеток с образованием многоядерных клеток, симпластов и синцитиев. Вирусы вызывают два типа слияния клеток: 1) «слияние снаружи» и 2) «слияние изнутри». «Слияние снаружи» происходит при высокой множественности инфекции и обнаруживается в течение первых часов после заражения. Этот тип слияния, описанный для парамиксовирусов, обусловлен белками заражающего вируса и не требует внутриклеточного синтеза вирусных компонентов. Напротив, «слияние изнутри» происходит при низкой множественности инфекции, обнаруживается на сравнительно поздних стадиях инфекционного процесса и обусловлено вновь синтезированными вирусными белками (рис. 9). «Слияние изнутри» описано для многих вирусов: вирусов герпеса, онковирусов, возбудителей медленных инфекций и др. Этот тип слияния вызывают те же вирусные гликопротеиды, которые обеспечивают проникновение вируса в клетку.
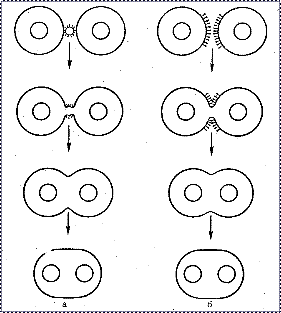 |
Рис. 9. Слияние клеточных мембран при заражении клеток вирусом:
а - слияние извне; б - слияние изнутри.
РАЗДЕВАНИЕ ВИРУСА В КЛЕТКЕ
Проникшие в клетку вирусные частицы должны раздеться для того, чтобы вызвать инфекционный процесс. Смысл раздевания заключается в удалении вирусных защитных оболочек, которые препятствуют экспрессии вирусного генома. В результате раздевания освобождается внутренний компонент вируса, который способен вызвать инфекционный процесс. Раздевание сопровождается рядом характерных особенностей: в результате распада вирусной частицы исчезает инфекционная активность, в ряде случаев появляется чувствительность к нуклеазам, возникает устойчивость к нейтрализующему действию антител, теряется фоточувствительность при использовании ряда препаратов.
Конечными продуктами раздевания являются сердцевины, нуклеокапсиды или нуклеиновые кислоты. Для ряда вирусов было показано, что продуктом раздевания являются не голые нуклеиновые кислоты, а нуклеиновые кислоты, связанные с внутренним вирусным белком. Например, конечным продуктом раздевания пикорнавирусов является РНК, ковалентно связанная с белком VРg, конечным продуктом раздевания аденовирусов является ДНК, ковалентно связанная с одним из внутренних вирусных белков.
В ряде случаев способность вирусов вызвать инфекционный процесс определяется возможностью их раздевания в клетке данной системы. Тем самым эта стадия является одной из стадий, лимитирующих инфекцию.
Раздевание ряда вирусов происходит в специализированных участках внутри клетки (лизосомах, структурах аппарата Гольджи, околоядерном пространстве, ядерных порах на ядерной мембране). При слиянии вирусной и клеточной мембран проникновение в клетку сочетается с раздеванием.
Раздевание и внутриклеточный транспорт являются взаимосвязанными процессами: при нарушении правильного внутриклеточного транспорта к местам раздевания вирусная частица попадает в лизосому и разрушается лизосомальными ферментами.








