2014-02-05
2014-02-05 1143
1143ТРАНСКРИПЦИЯ
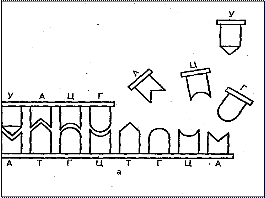 |
Транскрипция — это переписывание ДНК на РНК по законам генетического кода. Это означает, что РНК состоит из нуклеотидных последовательностей, комплементарных ДНК. Нити ДНК в участке транскрипции разделяются и функционируют как матрицы, к которым присоединяются комплементарные нуклеотиды благодаря спариванию комплементарных оснований (аденин связывается с тимином, урацил — с аденином, гуанин — с цитозином и цитозин — с гуанином) (рис. 10).
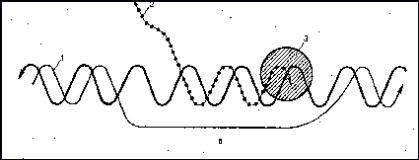 |
Рис. 10. Транскрипция ДНК и образование комплементарной РНК-цепочки:
а - спаривание комплементарных нуклеотидов при полимеризации;
А, Ц, Г, Т, У- сокращенные обозначения аденина, цитозина, гуанина, тимина, урацила;
б - схема транскрипции ДНК:
1 - ДНК; 2 - растущая нить РНК; 3 - ДНК-зависимая РНК-полимераза.
Транскрипция осуществляется с помощью специального фермента — РНК-полимеразы, который связывает нуклеотиды путем образования 3-5´фосфодиэфирных мостиков. Такое связывание происходит лишь в присутствии ДНК-матрицы.
Продуктами транскрипции в клетке являются иРНК. Сама клеточная ДНК, являющаяся носителем генетической информации, не может непосредственно программировать синтез белка. Передачу генетической информации от ДНК к рибосомам осуществляет РНК-посредник. На этом основана центральная догма молекулярной биологии, которая выражается следующей формулой:
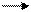
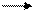
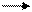 ДНК транскрипция РНК трансляция белок,
ДНК транскрипция РНК трансляция белок,
где стрелки показывают направление переноса генетической информации.
Реализация генетической информации у вирусов. Стратегия вирусного генома в отношении синтеза иРНК у разных вирусов различна. У ДНК-содержащих вирусов иРНК синтезируется на матрице одной из нитей ДНК. Формула переноса генетической информации у них такая же, как и в клетке:
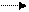
 ДНК транскрипция РНК трансляция белок.
ДНК транскрипция РНК трансляция белок.
ДНК-содержащие вирусы, репродукция которых происходит в ядре, используют для транскрипции клеточную полимеразу. К этим вирусам относятся паповавирусы, аденовирусы, вирусы герпеса. ДНК-содержащие вирусы, репродукция которых происходит в цитоплазме, не могут использовать клеточный фермент, находящийся в ядре. Транскрипция их генома осуществляется вирусспецифическим ферментом — ДНК-полимеразой, которая проникает в клетку в составе вируса. К этим вирусам относятся вирусы оспы и иридовирусы.
РНК-содержащие вирусы, у которых хранителем генетической информации является не ДНК, а РНК, решают эту проблему особым образом. У РНК-содержащих «плюс»-нитевых вирусов, у которых функции иРНК выполняет сам геном, передача генетической информации осуществляется по наиболее простой формуле:
 РНК белок.
РНК белок.
К этой группе вирусов относятся пикорнавирусы, тогавирусы, коронавирусы. У них нет необходимости в акте транскрипции для синтеза вирусспецифических белков. Поэтому транскрипцию как самостоятельный процесс у этих вирусов не выделяют.
Иначе дело обстоит у вирусов, геном которых не может выполнять функцию иРНК. В клетке синтезируется комплементарная геному РНК, которая и является информационной. Передача генетической информации у этих вирусов осуществляется по формуле:
 РНК - РНК белок
РНК - РНК белок
У этих вирусов транскрипция выделена как самостоятельный процесс в инфекционном цикле. К ним относятся две группы вирусов животных.
1. Вирусы, геном которых представлен однонитчатой РНК: ортомиксовирусы, парамиксовирусы, рабдовирусы, буньявирусы. Поскольку геномная РНК этих вирусов является «минус»-нитью, указанную группу вирусов называют «минус»-нитевыми вирусами.
2. Вирусы, геном которых представлен двунитчатой РНК (диплоРНК-вирусы). Среди вирусов животных к ним относятся реовирусы.
В клетке нет фермента, который может полимеризовать нуклеотиды на матрицеРНК. Эту функцию выполняет вирусспецифический фермент — РНК-полимераза, или транскриптаза, которая находится в составе вирусов и вместе с ними проникает в клетку.
Среди РНК-содержащих вирусов животных есть семейство ретровирусов, которые имеют уникальный путь передачи генетической информации. РНК этих вирусов переписывается на ДНК, ДНК интегрирует с клеточным геномом и в его составе переписывается на РНК, которая обладает информационными функциями. Путь передачи генетической информации в этом случае осуществляется по более сложной формуле:
 РНК - ДНК РНК белок
РНК - ДНК РНК белок
В составе этих вирусов есть уникальный вирусспецифический фермент, который переписывает РНК на ДНК. Этот процесс называется обратной транскрипцией, а фермент — обратная транскриптаза, или ревертаза. Тот же фермент синтезирует нить ДНК на матрице ДНК. Двунитчатая ДНК после замыкания в кольцо интегрирует с клеточным геномом, и транскрипцию интегрированной ДНК в составе клеточных геномов осуществляет клеточная РНК-полимераза. Поскольку иРНК ретровирусов гомологична геномной РНК (а не комплементарна ей), ретровирусы являются «плюс»-нитевыми вирусами.
Ферменты, транскрибирующие вирусный геном. Транскрипция ряда ДНК-содержащих вирусов — паповавирусов, аденовирусов, вирусов герпеса, парвовирусов, гепаднавирусов Осуществляется в ядре клетки, и в этом процессе широко используются механизмы клеточной транскрипции — ферменты транскрипции и дальнейшей модификации транскриптов. Транскрипция этих вирусов осуществляется клеточной РНК-полимеразой II — ферментом, который осуществляет транскрипцию клеточного генома. Однако особая группа транскриптов аденовируса синтезируется с помощью другого клеточного фермента — РНК-полимеразы III. У двух других семейств ДНК-содержащих вирусов животных — вирусов оспы и иридовирусов — транскрипция происходит в цитоплазме. Поскольку в цитоплазме нет клеточных полимераз, транскрипция этих вирусов нуждается в специальном вирусном ферменте — вирусной РНК-полимеразе. Этот фермент является структурным вирусным белком.
У РНК-содержащих вирусов транскрипция осуществляется вирус-специфическими транскриптазами, т. е. ферментами, закодированными в вирусном геноме. Вирусспецифические транскриптазы могут быть как структурными белками, входящими в состав вириона (эндогенная транскриптаза), так и неструктурными белками, которые синтезируются в зараженной клетке, но не включаются в вирион.
Транскрипция в зараженной клетке. Синтез комплементарных РНК на родительских матрицах с помощью родительской транскриптазы носит название первичной транскрипции в отличие от вторичной транскрипции, происходящей на более поздних стадиях инфекционного цикла на вновь синтезированных, дочерних матрицах, с помощью вновь синтезированной транскриптазы. Большая часть иРНК в зараженной клетке является продуктом вторичной транскрипции.
Транскриптивные комплексы. У сложно устроенных РНК-содержащих вирусов животных транскрипция происходит не на матрице голой РНК, а в составе вирусных нуклеокапсидов или сердцевин (транскриптивные комплексы). Связанные с геномом капсидные белки не только не препятствуют транскрипции, но и необходимы для нее, обеспечивая правильную конформацию тяжа РНК, защиту его от клеточных протеаз, связь отдельных фрагментов генома друг с другом, а также регуляцию транскрипции.
Вновь синтезированные иРНК выходят из транскриптивных комплексов и транспортируются к рибосомам.
На модели реовирусов было показано, что обе нити двунитчатых молекул РНК остаются в составе сердцевины, а вновь синтезированные иРНК выталкиваются из сердцевины через отверстия в 12 полых цилиндров, находящихся в составе сердцевины (рис. 11).
 |
Рис. 11. Вытеснение вновь синтезированной РНК из сердцевины вирионов реовирусов. Вид в электронном микроскопе (а) и схема строения вирионов (б).
Регуляция транскрипции. Транскрипция вирусного генома строго регулируется на протяжении инфекционного цикла. Регуляция осуществляется как клеточными, так и вирусспецифическими механизмами. У некоторых вирусов, в основном ДНК-содержащих, существует три периода транскрипций — сверхранняя, ранняя и поздняя. К этим вирусам относятся вирусы оспы, герпеса, паповавирусы, аденовирусы. В результате сверхранней и ранней транскрипции избирательно считываются сверхранние и ранние гены с образованием сверхранних или ранних иРНК. При поздней транскрипции считывается другая часть вирусного генома — поздние гены, с образованием поздних иРНК. Количество поздних генов обычно превышает количество ранних генов. Многие сверхранние гены являются генами для неструктурных белков — ферментов и регуляторов транскрипции и репликации вирусного генома. Напротив, поздние гены обычно являются генами для структурных белков. Обычно при поздней транскрипции считывается весь геном, но с преобладанием транскрипции поздних генов.
Фактором регуляции транскрипции у ядерных вирусов является транспорт транскриптов из ядра в цитоплазму, к месту функционирования иРНК — полисомам.
Продуктом сверхранней транскрипции вирусов герпеса являются А-белки. Функция одного или нескольких из них необходима для транскрипции следующей группы генов, кодирующих Р-белки. В свою очередь Р-белки включают транскрипцию последней группы поздних генов, кодирующих У-белки. Такой тип регуляции получил название «каскадной».
У РНК-содержащих вирусов синтез транскриптов также строго контролируется в отношении как количества каждого класса транскриптов, так и периода инфекции, когда определенные транскрипты синтезируются с максимальной скоростью. На ранней стадии инфекции преимущественно синтезируются транскрипты двух генов вируса гриппа — NР и NS, на поздней стадии инфекции — транскрипты генов М, НА и NА. Остальные три гена для Р-белков синтезируются примерно с одинаковой скоростью на протяжении всего периода инфекции. У реовирусов на ранней стадии инфекции преимущественно транскрибируется 4 из 10 фрагментов генома и лишь на поздней стадии транскрибируется весь геном. Однако если поместить геном вируса в бесклеточную РНК-синтезирующую систему, будет происходить равномерная транскрипция всех 10 фрагментов генома. Эти факты говорят о жестком контроле транскрипции со стороны клетки-хозяина и возможном наличии специфических клеточных регуляторов.
У парамиксовирусов и рабдовирусов весь геном представляет собой одну транскрипционную единицу с единственным промотором (участок связывания транскриптазы и начала транскрипции) у 3'-конца. Вдоль генома существует как бы градиент эффективности транскрипции. Ближайший к 3'- концу ген (ген наиболее обильного белка КР) считывается наиболее часто. Напротив, ген для самого высокомолекулярного белка — транскриптазы,— содержащегося лишь в количестве нескольких молекул на вирион, находится на противоположном конце генома и транскрибируется значительно реже. Такая регуляция экспрессии генов путем порядка их расположения в геноме носит название «полярность». При этом способе регуляции количество молекул полипептидов определяется полярностью гена, т.е. расстоянием его от промотора.
ТРАНСЛЯЦИЯ
Синтез белка в клетке происходит в результате трансляции иРНК. Трансляцией называется процесс перевода генетической информации, содержащейся в иРНК, на специфическую последовательность аминокислот. Иными словами, в процессе трансляции осуществляется перевод 4-буквенного языка азотистых оснований на 20-буквенный язык аминокислот.
ТранспортныеРНК. Свою аминокислоту тРНК узнают по конфигурации ее боковой цепи, а специфический фермент аминоацил-синтетаза катализирует ассоциацию тРНК с аминокислотой. В клетке существует большое количество разнообразных видов тРНК. Поскольку для каждой аминокислоты должна быть своя тРНК, количество видов тРНК должно быть не меньше 20, однако в клетке их значительно больше. Это связано с тем, что для каждой аминокислоты существует не один, а несколько видов тРНК. Молекула тРНК представляет собой однонитчатую РНК со сложной структурой в виде кленового листа (рис. 12). Один ее конец связывается с аминокислотой (конец а), а противоположный — с нуклеотидами иРНК, которым они комплементарны (конец б). Три нуклеотида на иРНК кодируют одну аминокислоту и называются «триплет» или «кодон», комплементарные кодону три нуклеотида на конце тРНК называются «антикодон».
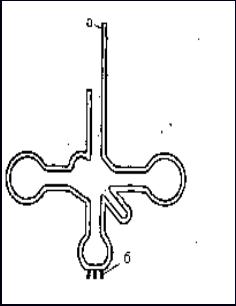 |
Рис. 12. Строение транспортной РНК:
а - участок связывания с аминокислотой;
б - участок связывания с иРНК (антикодон).
Рибосомы. Синтез белка в клетке осуществляется на рибосоме. Рибосома состоит из двух субъединиц, большой и малой, малая субъединица примерно в два раза меньше большой. Обе субъединицы содержат по одной молекуле рибосомальной РНК и ряд белков. Рибосомальные РНК синтезируются в ядре на матрице ДНК с помощью РНК-полимеразы I. В малой рибосомальной субъединице есть канал, в котором находится информационная РНК. В большой рибосомальной субъединице есть две полости, захватывающие также малую рибосомальную субъединицу. Одна из них содержит аминоацильный центр (А-центр), другая — пептидильный центр (П-центр) (рис. 13).
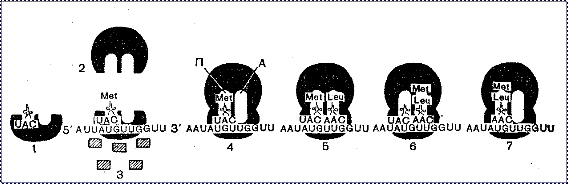
Рис. 13. Формирование и функционирование рибосомы:
1 - малая рибосомальная субъединица с присоединенной инициаторной метионил-тРНК;
2 - большая рибосомальная субъединица;
3 - инициаторный комплекс, содержащий малую рибосомальную субъединицу, метионил-тРНК и иРНК;
заштрихованные прямоугольники - белковые факторы инициации (9 факторов в эукариотических клетках);
4 - функционально активная рибосома: А — аминоацильный центр, П— пептидильный центр в большой рибосомальной субъединице;
5, 6, 7 - процесс элонгации полипептидной цепи; показан перенос аминоацил-тРНК между двумя центрами на большой рибосомальной субъединице, осуществляемый с помощью пептидил-трансферазы.
Фазы трансляции. Процесс трансляции состоит из трех фаз: 1) инициации, 2) элонгации и 3) терминации.
Инициация трансляции. Это наиболее ответственный этап в процессе трансляции, основанный на узнавании рибосомой иРНК и связывании с ее особыми участками. Рибосома узнает иРНК благодаря «шапочке» на 5'-конце и скользит к 3'-концу, пока не достигнет инициаторного кодона, с которого начинается трансляция. В эукариотической клетке инициаторным кодоном является кодон АУГ или ГУГ, кодирующие метионин. С метионина начинается синтез всех полипептидных цепей.
В начале с иРНК связывается малая рибосомальная субъединица. К комплексу иРНК с малой рибосомальной субъединицей присоединяются другие компоненты, необходимые для начала трансляции. Это несколько молекул белка, которые называются «инициаторные факторы». Их по крайней мере три в прокариотической клетке и более девяти в эукариотической клетке.
Инициаторные факторы определяют узнавание рибосомой специфических иРНК и, таким образом, являются определяющим фактором в дискриминации между различными иРНК, присутствующими в клетке, как правило, в избыточном количестве.
В результате формируется комплекс, необходимый для инициации трансляции, который называется инициаторным комплексом. В инициаторный комплекс входят:
1) иРНК;
2) малая рибосомальная субъединица;
3) аминоацил-тРНК, несущая инициаторную аминокислоту;
4) инициаторные факторы;
5) несколько молекулГТФ.
В рибосоме осуществляется слияние потока информации с потоком аминокислот. Аминоацил-тРНК входит в А-центр большой рибосомальной субъединицы, и ее антикодон взаимодействует с кодоном иРНК, находящейся в малой рибосомальной субъединице. При продвижении иРНК на один кодон тРНК перебрасывается в пептидильный центр, и ее аминокислота присоединяется к инициаторной аминокислоте с образованием первой пептидной связи. Свободная от аминокислоты тРНК, выходит из рибосомы, и может опять функционировать в транспорте специфических аминокислот. На ее место из А-центра в П-центр перебрасывается новая тРНК и образуется новая пептидная связь. В А-центре появляется вакантный кодон иРНК, к которому немедленно присоединяется соответствующая тРНК и происходит присоединение новых аминокислот к растущей полипептидной цепи (рис. 13).
Элонгация трансляции. Это процесс удлинения, наращивания полипептидной цепи, основанный на присоединении новых аминокислот с помощью пептидной связи. Происходит постоянное протягивание нити иРНК через рибосому и «декодирование» заложенной в ней генетической информации (рис. 14). иРНК функционирует на нескольких рибосомах, каждая из которых синтезирует одну и ту же полипептидную нить, кодируемую данной иРНК. Группа рибосом, работающих на одной молекуле иРНК, называется полирибосомой, или полисомой. Размер полисом значительно варьирует в зависимости от длины молекулы иРНК, а также от расстояния между рибосомами. Так, полисомы, которые синтезируют гемоглобин, состоят из 4-6 рибосом, высокомолекулярные белки синтезируются на полирибосомах, содержащих 20 и более рибосом.
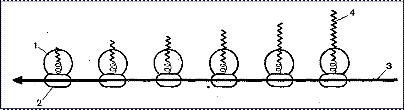 |
Рис. 14. Синтез белков на полисомах:
1 - большая рибосомальная субъединица;
2 - малая рибосомальная субъединица;
3 - иРНК;
4 - растущая полипептидная нить.
Терминация трансляции. Терминация трансляции происходит в тот момент, когда рибосома доходит до терминирующего кодона в составе иРНК. Трансляция прекращается, и полипептидная цепь освобождается из полирибосомы. После окончания трансляции полирибосомы распадаются на субъединицы, которые могут войти в состав новых полирибосом.
Свойства полирибосом. По топографии в клетке полирибосомы делят на две большие группы — свободные и связанные с мембранами эндоплазматической сети, которые составляют соответственно 75 и 25 %.
Между двумя группами полирибосом нет принципиальных структурных и функциональных различий, они формируются из одного и того же пула субъединиц и в процессе трансляции могут обмениваться субъединицами. Мембраны, с которыми связаны полирибосомы, называются грубыми или шероховатыми мембранами в отличие от гладких мембран, не содержащих полирибосомы. Связь полирибосом с мембранами осуществляется с помощью сигнального пептида — специфической последовательности на аминоконце синтезирующихся гликопротеидов. На связанных с мембранами полирибосомах синтезируются внутримембранные белки, которые, сразу же после синтеза, оказываются в составе мембран.
Трансляция в зараженных вирусом клетках. Стратегия вирусного генома, использующего клеточный аппарат трансляции, должна быть направлена на создание механизма для подавления трансляции собственных клеточных иРНК и для избирательной трансляции вирусных иРНК, которые всегда находятся в значительно меньшем количестве, чем клеточные матрицы. Этот механизм реализуется на уровне специфического узнавания малой рибосомальной субъединицей вирусных иРНК, т. е. на уровне формирования инициирующего комплекса. Поскольку многие вирусы не подавляют синтез клеточных иРНК, в зараженных клетках возникает парадоксальная ситуация: прекращается трансляция огромного фонда функционально активных клеточных иРНК, и на освободившихся рибосомах начинается трансляция одиночных молекул вирусных иРНК. Специфическое узнавание рибосомой вирусных иРНК осуществляется за счет вирусспецифических инициаторных факторов.
Два способа формирования вирусных белков. Поскольку геном вируса животных представлен молекулой, кодирующей более чем один белок, вирусы поставлены перед необходимостью синтеза либо длинной иРНК, кодирующей один гигантский полипептид-предшественник, который затем должен быть нарезан в специфических точках на функционально активные белки, либо коротких моноцистронных иРНК, каждая из которых кодирует один белок.
Таким образом, существуют два способа формирования вирусных белков: 1) иРНК транслируется в гигантский полипептид-предшественник, который после синтеза последовательно нарезается на зрелые функционально активные белки; 2) иРНК транслируется с образованием зрелых белков, или белков, которые лишь незначительно модифицируются после синтеза.
Первый способ трансляции характерен для РНК-содержащих «плюс»-нитевых вирусов — пикорнавирусов и тогавирусов. Их иРНК транслируется в гигантскую полипептидную цепь, так называемый полипротеид, который сползает в виде непрерывной ленты с рибосомного «конвейера» и нарезается на индивидуальные белки нужного размера. Нарезание вирусных белков является многоступенчатым процессом, осуществляемым как вирусспецифическими, так и клеточными протеазами. В клетках, зараженных пикорнавирусами, на конце полипротеина-предшественника находится белок с протеазной активностью. Вирусная протеаза осуществляет нарезание предшественника на 3 фрагмента, один из которых является предшественником для структурных белков, второй — для неструктурных белков, функции третьего фрагмента неизвестны. В дальнейшем нарезании участвуют вирус-специфические и клеточные протеазы.
Интересный вариант первого способа трансляции обнаруживается у альфа-вирусов. Геномная РНК с коэффициентом седиментации 42 S транслируется с образованием полипептида-предшественника для неструктурных белков. Однако доминирующей в зараженных клетках иРНК является РНК с коэффициентом седиментации 26 S, составляющая одну треть геномной РНК. Эта иРНК транслируется с образованием предшественника для структурных белков.
Второй способ формирования белков характерен для ДНК-содержащих вирусов и большинства РНК-содержащих вирусов. При этом способе синтезируются короткие моноцистронные иРНК в результате избирательной транскрипции одного участка генома (гена). Однако все вирусы широко используют механизм посттрансляционного нарезания белка.
Вирусспецифические полисомы. Поскольку длина вирусныхиРНК варьирует в широких пределах, размер вирусспецифических полисом также широко варьирует: от 3-4 до нескольких десятков рибосом на одной нити иРНК. При инфекциях, вызванных пикорнавирусами, формируются крупные полисомы, представляющие собой агрегаты, состоящие из 20-60 рибосом. При инфекциях, вызванных другими вирусами животных, использующими второй способ трансляции, формируются полисомы небольшого размера. Между размерами иРНК и величиной полисом существует определенная корреляция, однако в ряде Случаев полисомы имеют больший или меньший размер по сравнению с ожидаемым. Эта особенность вирусных полисом объясняется необычным пространственным расположением рибосом на вирусных матрицах, связанных с меньшей плотностью упаковки рибосом на молекулеиРНК.
Вирусспецифические полисомы могут быть как свободными, так и связанными с мембранами. В зараженных вирусом полиомиелита клетках полипротеид синтезируется на связанных с мембранами полисомах; при инфекциях, вызванных сложно устроенными вирусами, формируются как свободные, так и связанные с мембранами полисомы, которые вовлечены в синтез разных классов вирусных полипептидов. Внутренние белки обычно синтезируются на свободных полисомах, гликопротеиды всегда синтезируются на полисомах, связанных с мембранами.
Модификация вирусных белков. В эукариотической клетке многие белки, в том числе вирусные, подвергаются посттрансляционным модификациям, и зрелые функционально активные белки часто не идентичны их вновь синтезированным предшественникам. Широко распространены такие посттрансляционные ковалентные модификации, как гликозилирование, ацилирование, метилирование, сульфирование (образование дисульфидных связей), протеолитическое нарезание и, наконец, фосфорилирование. В результате вместо 20 генетически закодированных аминокислот из различных клеток разных органов эукариотов выделено около 140 дериватов аминокислот.
Среди широкого спектра модифицированных реакций лишь небольшое количество процессов является обратимыми:
1) фосфорилирование — дефосфорилирование;
2) ацилирование — деацилирование;
3) метилирование — деметилирование;
4) образование дисульфидных связей.
Среди подобных обратимых модификаций белков следует искать процессы, обусловливающие механизм регуляции активности белков в эукариотической клетке.
Гликозилирование. В составе сложно устроенных РНК- и ДНК-содержащих вирусов имеются белки, содержащие ковалентно присоединенные боковые цепочки углеводов — гликопротеиды. Гликопротеиды расположены в составе вирусных оболочек и находятся на поверхности вирусных частиц. Своей гидрофобной частью они погружены в двойной слой липидов, а некоторые гликопротеиды проникают через него и взаимодействуют с внутренним компонентом вируса. Гидрофильная часть молекулы обращена наружу.
Синтез и внутриклеточный транспорт гликопротеидов характеризуется рядом особенностей, присущих клеточным внутримембранным белкам. Их синтез осуществляется на полисомах, ассоциированных с мембранами, и белки сразу же после синтеза попадают в шероховатые мембраны, откуда транспортируются в мембраны эндоплазматической сети и в комплекс Гольджи, где происходит модификация и комплектование углеводной цепочки, а затем — в плазматическую мембрану в ряде случаев путем слияния с ней везикул комплекса Гольджи. Такой целенаправленный транспорт осуществляется благодаря имеющейся на аминоконце белка специфической последовательности из 20-30 аминокислот (сигнальному пептиду). Сигнальный пептид отрезается от белковой молекулы после того, как гликопротеид достигает плазматической мембраны.
Гликозилирование полипептидов является сложным многоступенчатым процессом, первые этапы которого начинаются уже в процессе синтеза полипептидов, и первый сахар присоединяется к полипептидной цепи, еще не сошедшей с рибосомы. Последующие этапы гликози-лирования происходят путем последовательного присоединения Сахаров в виде блоков к углеводной цепочке в процессе транспорта полипептида к плазматической мембране.
Окончательное формирование углеводной цепочки может завершаться на плазматической мембране перед сборкой вирусной частицы. Процесс гликозилирования не влияет на транспорт полипептида к плазматической мембране, но имеет существенное значение для экспрессии биологической активности белка. При подавлении гликозилирования соответствующими ингибиторами (аналоги сахаров типа 2-дезоксиглюкозы, антибиотик туникамицин) нарушается синтез полипептидов, блокируется сборка вирионов миксовирусов, рабдовирусов, альфа-вирусов или образуются неинфекционные вирионы герпеса и онковирусов.
Сульфирование. Некоторые белки сложно устроенных РНК- и ДНК-содержащих вирусов сульфируются после трансляции. Чаще всего сульфированию подвергаются гликопротеиды, при этом сульфатная группа связывается с сахарным компонентом гликопротеида.
Ацилирование. Ряд гликопротеидов сложно устроенных РНК-содержащих вирусов (НА2 вируса гриппа, белок G вируса везикулярного стоматита, белок НN вируса ньюкаслской болезни и др.) содержат ковалентно связанные 1-2 молекулы жирных кислот.
Нарезание. Многие вирусные белки ив первую очередь гликопротеиды приобретают функциональную активность лишь после того, как произойдет их нарезание в специфических точках протеолитическими ферментами. Нарезание происходит либо с образованием двух функциональных белковых субъединиц (например, большая и малая субъединицы гемагглютинина вируса гриппа, два гликопротеида, E2 и Е3, вируса леса Семлики) либо с образованием одного функционально активного белка и неактивного фрагмента, например белки F и HN парамиксовирусов. Нарезание обычно осуществляется клеточными ферментами. У многих сложно устроенных вирусов животных, имеющих гликопротеид, нарезание необходимо для формирования активных прикрепительных белков и белков слияния и, следовательно, для приобретения вирусом способности инфицировать клетку. Лишь после нарезания этих белков вирусная частица приобретает инфекционную активность. Таким образом, можно говорить о протеолитической активации ряда вирусов, осуществляемой с помощью клеточных ферментов.
Фосфорилирование. Фосфорпротеиды содержатся практически в составе всех вирусов животных, РНК- и ДНК-содержащих, просто и сложно устроенных. В составе большинства вирусов обнаружены протеинкиназы, однако фосфорилирование может осуществляться как вирусными, так и клеточными ферментами. Обычно фосфорилируются белки, связанные с вирусным геномом и осуществляющие регулирующую роль в его экспрессии.
С процессом фосфорилирования связан механизм антивирусного действия интерферона. В зараженных вирусом клетках интерферон индуцирует синтез протеинкиназы, которая фосфорилирует субъединицу инициирующего фактора трансляции ЭИФ-2, в результате чего блокируется трансляция вирусных информационных РНК. Фосфорилирование белков играет регулирующую роль в транскрипции и трансляции вирусных иРНК, специфическом узнавании вирусных иРНК рибосомой, белок-нуклеиновом и белок-белковом узнавании на стадии сборки вирусных частиц.