 2014-02-05
2014-02-05 6504
6504Таблица 8.
КЛАССИФИКАЦИЯ ВИРУСНЫХ ИНФЕКЦИЙ НА УРОВНЕ ОРГАНИЗМА
КЛАССИФИКАЦИЯ ВИРУСНЫХ ИНФЕКЦИЙ НА УРОВНЕ КЛЕТКИ
ГЛАВА 6. ПАТОГЕНЕЗ ВИРУСНЫХ ИНФЕКЦИЙ
ГЛАВА 5. ГЕНЕТИКА ВИРУСОВ
Величайшие достижения серединыXX века — открытие дискретных единиц наследственности (генов), разработка хромосомной теории наследственности, развитие биохимической генетики микроорганизмов и установление принципа «один ген — один белок», открытие регуляции активности генов прокариотов Ф. Жакобом и Ж. Моно, открытие двойной спирали ДНК Дж. Уотсоном и Ф. Криком и др., — создали основу для превращения генетики классической в генетику молекулярную, где законы наследственности и изменчивости изучаются на молекулярном и субмолекулярном уровнях.
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ГЕНОМА ВИРУСА
Вирусы являются одним из излюбленных объектов молекулярной генетики благодаря простому строению и малой молекулярной массеих геномов, которая в 106 раз меньше массы генома эукариотической клетки. Организация генетического аппарата у ряда вирусов, например у SV40, настолько сходна с таковой генов эукариотической клетки, что получила название минихромосомы. Минихромосома широко используется для изучения организации и репликации ДНК.
Число генов у вирусов значительно варьирует: от 3-4 генов у просто устроенных вирусов (парвовирусы) до150 генов и больше у сложно устроенных (вирус оспы). Геном вирусов животных является гаплоидным, за исключением ретровирусов, которые имеют диплоидный геном, представленный двумя идентичными молекулами РНК. У вирусов с фрагментарным геномом (вирусы гриппа, реовирусы) каждый фрагмент обычно представляет собой один ген.
Так же, как и геном эукариотической клетки, ДНК-геном ряда вирусов животных имеет мозаичную структуру, при которой смысловые последовательности чередуются с неинформативными последовательностями. Механизм сплайсинга при формировании иРНК широко распространен и среди вирусов, имеющих ядерную локализацию транскрипции (адено-, папова-, герпесвирусы), поскольку ферменты, осуществляющие сплайсинг, находятся в ядре. Однако сплайсинг был обнаружен и у РНК-содержащих вирусов. Например, у вирусов гриппа происходит сплайсинг транскриптов 7-го и 8-го генов; в результате сплайсинга и сдвига рамки трансляции продуктами каждого из этих генов являются по два уникальных белка.
В составе генов ДНК-содержащих вирусов есть регуляторные участки, в том числе промотор, контролирующие функцию структурных генов. Сильными промоторами являются концы многих вирусных ДНК, представляющие собой длинные концевые повторы, сильный промоторимеют гены тимидинкиназы вирусов оспы и герпеса. Эти промоторы используются в генной инженерии для усиления транскрипции изучаемого гена.
ГЕНЕТИЧЕСКИЕ И НЕГЕНЕТИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ ВИРУСОВ
Взаимодействия между вирусами возникают в условиях смешанной инфекции, когда два вируса или более, родственных или неродственных, заражают одну и ту же клетку. Различают генетические и негенетические взаимодействия вирусов.
К генетическим взаимодействиям относят только те, в результате которых происходит обмен генетического материала, и возникают геномы, содержащие фрагменты обоих вирусов-родителей. К генетическим взаимодействиям относят множественную реактивацию, рекомбинацию, кросс- реактивацию, реассортацию и гетерозиготность.
Множественнаяреактивация. Вирусная инфекция может возникнуть при заражении клетки несколькими вирионами с поврежденными геномами вследствие того, что функцию поврежденного гена может выполнять вирус, у которого этот ген не поврежден. Этот феномен был вначале обнаружен на бактериофагах и получил название множественной реактивации. В основе множественной реактивации лежит кооперативный процесс, при котором вирионы с поражением разных генов дополняют друг друга путем генетической рекомбинации, в результате чего репродуцируется исходный неповрежденный вирус.
Эффективность множественности реактивации зависит от многих причин: степени повреждения генома вирионов, числа проникших в клетку вирионов, концентрации их в определенных участках клетки, аутоинтерференции поврежденных вирионов. Для множественной реактивации важное значение имеет расстояние между вирионами с поврежденными геномами внутри клетки. Обработка вирионов двухвалентными ионами металлов, ведущая к их агрегации, усиливает множественную реактивацию.
Рекомбинацией называют обмен частями генома, включающей ковалентное встраивание участка (или участков) генома одного вируса в геном другого.
Под реассортацией понимают обмен геномными сегментами, когда не происходит ковалентного встраивания, и который возможен только у тех вирусов, чей геном представлен раздельными сегментами, каждый из которых включает один или несколько генов (вирусы с сегментированным геномом). Она наблюдается при генетических взаимодействиях между вирусами, имеющими сегментированный геном. Образующиеся при этом гибридные формы вирусов называют реассортантами. Реассортанты вирусов гриппа получают при совместном культивировании вирусов с разными генами гемагглютинина и нейраминидазы. В этом случае из общего потомства путем нейтрализации соответствующих антигенов можно выделить интересующие исследователя варианты.
Существуют определенные группировки (констелляцииили созвездия) генов, которые в данной системе клеток более стойки и делают вирус более жизнеспособным.
Сходные процессы пересортировки генов имеют место у вирусов гриппа типов А, В и С и у других вирусов с фрагментарным геном — у буньявирусов, аренавирусов (однонитчатые РНК) и реовирусов (ротавирусов) (двунитчатая РНК). Однако эти процессы не столь интенсивны и доступны изучению, как у вирусов гриппа.
Перекрестная реактивация. Перекрестная реактивация, кросс-реактивация или реактивация при скрещивании, происходит в том случае, когда у одного из штаммов вируса часть генома повреждена, а другой геном интактен. При смешанной инфекции двумя такими вирусами возможна рекомбинация неповрежденных участков генома инактивированного вируса с геномом интактного вируса, и в результате этого процесса появляются штаммы вируса со свойствами обоих родителей. Описываемый феномен также обозначается как «спасение маркера», поскольку реактивируется (рекомбинирует) лишь часть генома инактивированного вируса, несущая какой-нибудь признак (маркер).
Гетерозиготность. При совместном культивировании двух штаммов вируса может происходить формирование вирионов, содержащих в своем составе два разных генома или по крайней мере один полный геном и часть второго генома. Это явление названо гетерозиготностью.
Негенетическими взаимодействиями называют взаимодействия между белками разных вирусов (или с участием белков одного вируса и генетического материала другого вируса), не ведущие к наследуемым изменениям. При смешанной инфекции имеют место, как правило, и генетические и негенетические взаимодействия, причем последние могут принимать участие в генерации наследуемых перестроек генома, т.е. в генетических взаимодействиях. К негенетическим взаимодействиям вирусов относят комплементацию, фенотипическое смешивание и интерференцию.
Комплеменатацией называют взаимодействие белков разных вирусов в зараженной клетке, а также взаимодействие белков одного вируса с генетическим материалом (ДНК или РНК) другого, в результате которого репродукция вируса усиливается. Если ген одного из вирусов имеет мутацию, в результате которой соответствующий белок нефункционален, аналогичный белок другого вируса может восполнить недостающую функцию. Комплементация может быть неаллельной, межгенной, если каждый из двух вирусов имеет мутацию, которая не позволяет ему репродуцироваться в непермиссивных условиях, но мутации локализованы в двух разных генах, так что каждый вирус продуктом своего нормального, не имеющего мутации гена помогает вирусу-партнеру эффективно репродуцироваться. Возможна и неаллельная, внутригенная комплементация, в тех случаях, когда мутации у двух вирусов находятся в одном и том же гене, но в разных его участках, соответствующих разным доменам молекулы белка, кодируемого этим геном. Такая комплементация наблюдается в тех случаях, когда белок осуществляет свою функцию в виде олиго- или мультимера. Оба случая имеют место при комплементации между близкородственными вирусами. Комплементация возможна и между неродственными вирусами: например, вирусы-сателлиты (дельтавирус, аденоассоциированные вирусы и т.п.) способны к репродукции лишь в присутствии вируса-помощника.
Фенотипическим смешиванием называют формирование вириона, содержащего структурные белки двух вирусов. Вирусы, участвующие в фенотипическом смешивании, могут быть как близкородственными, так и неродственными. При фенотипическом смешивании гетерогенными, т.е. происходящими от разных вирусов, могут быть пепломеры в липопротеидной оболочке или капсомеры в капсиде. Частный случай фенотипического смешивания – транскапсидация, при которой геном одного вируса оказывается заключенным в капсид другого.
Интерференцией называют подавление размножения одного вируса другим. К негенетическим взаимодействиям обычно относят не опосредованную интерференцию (через индукцию интерферона или других факторов), а только ту, которая возникает при непосредственном взаимодействии вирусов, репродуцирующихся в одной клетке. К таким случаям относится подавление репродукции вируса дикого типа мутантом или холодо-адаптированным вариантом.
ОСНОВНЫЕ ПРОЦЕССЫ, КОНТРОЛИРУЮЩИЕ НАСЛЕДСТВЕННОСТЬ И ИЗМЕНЧИВОСТЬ ВИРУСОВ
Модификации. Модификациями называются не наследуемые (фенотипические) изменения у вирусов, обусловленные клеткой-хозяином. Эти изменения лежат в основе адаптации вируса к новому хозяину и преодоления зависимого от хозяина ограничения. Модификации нуклеиновых кислот вирусов осуществляют клеточные ферменты, ответственные за ограничение (рестрикцию) репродукции вируса.
Мутации. В основе изменчивости вирусов лежат мутации, т. е. изменения состава и последовательностей нуклеотидов вирусного генома. Мутации происходят у всех вирусов, независимо от того, является ли их генетическим аппаратом ДНК или РНК. В результате мутаций отдельные вирионы могут приобретать новые свойства. Дальнейшая судьба таких вирусов зависит от естественного отбора, сохраняющего популяцию, наиболее приспособленную к условиям существования. Мутации могут иметь разные последствия. В одних случаях они ведут к изменению фенотипических проявлений в нормальных условиях. Например, увеличивается или уменьшается размер бляшек под агаровым покрытием; увеличивается или ослабляется нейровирулентность для определенного вида животных; вирус становится более чувствительным к действию химиотерапевтического агента и т. п. В других случаях мутация является летальной, так как вследствие ее нарушается синтез или функция жизненно важного вирусспецифического белка, например вирусной полимеразы.
В некоторых случаях мутации являются условно летальными, так как вирусспецифический белок сохраняет свои функции в определенных, оптимальных для него, условиях и теряет эту способность в неразрешающих (непермиссивных) условиях. Типичным примером таких мутаций являются температурно-чувствительные (temperature sensitive) — ts-мутации, при которых вирус теряет способность размножения при повышенных температурах (39-42 °С), сохраняя эту способность при обычных температурах выращивания (36-37 °С).
По своему механизму мутации могут быть тоже разными. В одних случаях происходит деления, т. е. выпадение одного или нескольких нуклеотидов, в других случаях происходит встраивание одного или нескольких нуклеотидов, а в некоторых случаях — замена одного нуклеотида другим. Мутации могут быть прямыми и обратными. Прямые мутации меняют фенотип, а обратные мутации — реверсии — его восстанавливают. Возможны истинные реверсии, когда обратная мутация происходит в месте первичного повреждения, и псевдореверсии, если мутация происходит в другом участке дефектного гена (интрагенная супрессия) или в другом гене (экстрагенная супрессия). Реверсия не является редким событием, так как ревертанты обычно более приспособлены к данной клеточной системе. Поэтому при получении мутантов с заданными свойствами, например, вакцинных штаммов, приходится считаться с возможной их реверсией к дикому типу. Мутации носят случайный характер и объясняются статистическими законами.
В качестве физических мутагенов наиболее часто применяется ультрафиолетовое облучение, так как его энергия сопоставима с энергией химических связей. Реже применяются более жесткие виды облучения — рентгеновское и гамма-облучение, а также обработка вирусных суспензий нейтронами, протонами, электронами и ядрами гелия, так как они вызывают сильные разрушения вирусных геномов и их инактивацию.
В качестве химических мутагенов применяют аналоги оснований (бромурацил, бромдезоксиуридин, 2-аминопурин, нитрозогуанидин и пр.), алкилирующие и флуоресцирующие соединения (профлавин), интеркалирующие агенты (актиномицин, этидин бромид), азотистую кислоту, гидроксиламин и многие другие.
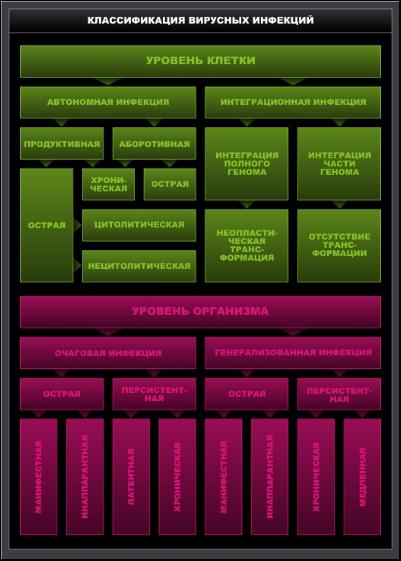 |
Под инфекцией понимают комплекс процессов, происходящих при взаимодействии инфекционного агента с организмом хозяина. Однако в связи с тем, что вирусы являются внутриклеточными паразитами, а точнее, генетическими паразитами, в основе их взаимодействия с организмом всегда лежит инфекционный процесс на уровне клетки, который реализуется путем взаимодействия вирусного и клеточного геномов. Поэтому возможно классифицировать инфекции как на клеточном уровне, так и на уровне организма (схема 2).
Схема 2. Классификация вирусных инфекций.
Автономные и интеграционные инфекции. Если вирусный геном реплицируется независимо от клеточного генома, такая инфекция называется автономной. Понятие автономии относительно, оно ограничивается лишь отсутствием физической связи между вирусным и клеточным геномами, хотя взаимодействие их постоянно происходит в течение инфекции. Автономная форма вирусной инфекции характерна для большинства вирусов животных.
Если вирусный геном включается в состав клеточного генома, или, как принято называть этот процесс, интегрирует с клеточным геномом и реплицируется вместе с ним, такая инфекция называется интеграционной. Интеграционная инфекция возникает в результате физического объединения генома вируса и клетки. При этой форме инфекции вирусный геном реплицируется и функционирует как составная часть клеточного генома. Интегрировать могут как полный геном, так и часть генома. При гепатите В возможна интеграция полного генома, при аденовирусных и герпесвирусных инфекциях обычно интегрирует часть генома, при инфекции онковирусами может интегрировать как полный геном, так и часть его. Вирусные последовательности в составе клеточного генома называются провирусом, или провирусной ДНК.
При интеграционных инфекциях нет ни сборки вирусной частицы, ни выхода вируса из клетки. Клетка может сохранить нормальные функции и при ее делении вирусные последовательности могут переходить в геном дочерних клеток. Такая ситуация наблюдается в случае инфекции, вызванной онкогенными вирусами. Интеграция может привести к неопластической трансформации клетки. Трансформированная клетка приобретает способность к неограниченному делению в результате нарушения регуляторных механизмов, контролирующих деление. Интеграционный тип инфекции возможен для нескольких семейств ДНК-содержащих вирусов: аденовирусов, паповавирусов, вирусов герпеса, а также для вируса гепатита В и обязателен для одного семейства РНК-содержащих вирусов — ретровирусов. В соответствии с данными В.М. Жданова, интеграционная форма инфекции может возникнуть при заражении и другими РНК-содержащими вирусами, такими, как вирус клещевого энцефалита (семейство тогавирусов), вирусы кори и SV5 (семейство парамиксовирусов) и др. Обязательным условием в этом случае является присутствие в клетках фермента — обратной транскриптазы, необходимого для процесса интеграции. Возникающая интеграционная инфекция может явиться причиной ряда хронических и аутоиммунных заболеваний.
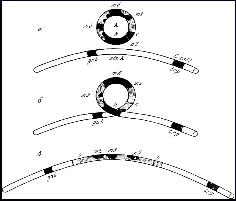 |
Механизм интеграции вирусного генома с клеточным геномом. Из многих моделей, объясняющих процесс интеграции, наиболее признанной является модель Кемпбелла. В соответствии с этой моделью для интеграции с клеточным геномом необходима кольцевая форма двунитчатой вирусной ДНК. Эта молекула ДНК прикрепляется к клеточной ДНК, в месте прикрепления обе молекулы разрезаются и образовавшиеся концы сшиваются таким образом, что вирусная ДНК становится частью клеточного генома (рис. 18).
Рис. 18. Модель Кэмпбелла (1962) интеграции профага:
а - кольцевая хромосома фага образует синапс с местом прикрепления фага λ (attλ) на хромосоме бактерии;
б - хромосома фага разрывается между генами h и c (в области b2), а хромосома бактерии разрывается между генами gal и trp;
в - в результате кроссинговера образуется одна непрерывная генетическая структура, содержащая геном фага между генами gal и trp.
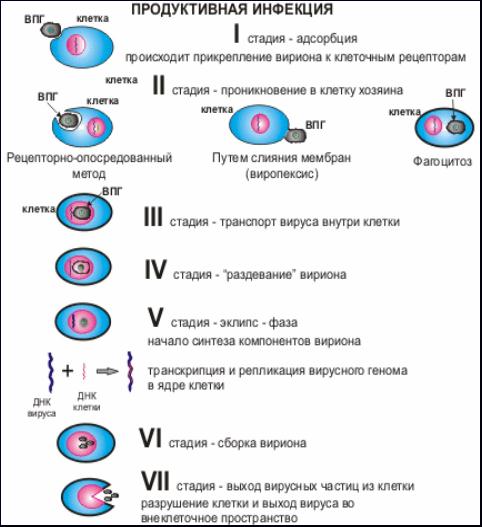 |
Продуктивная и интегративная формы инфекции. Инфекция может быть продуктивной и интегративной. Продуктивная инфекция завершается образованием инфекционного потомства (рис. 19). Этот тип инфекции возможен при наличии в зараженном организме пермиссивных (чувствительных) клеток, где реализуется репликтивный цикл возбудителя. При продуктивной вирусной инфекции происходит репродукция вирусов, а клетка погибает. Эта инфекция лежит в основе острых вирусных заболеваний, а так же в основе условных латентных инфекций, при которой погибают не все клетки пораженного органа, а только их часть. Неповрежденные клетки этого органа могут компенсировать временную утрату его функции, в следстии чего заболевание некоторое время е проявляеться, пока не наступит декомпенсация.
Рис.19. Продуктивная инфекция.
Интегративная форма инфекция (вирогения) – многие вирусы способны интегрировать свои нуклеиновые кислоты в геном клетки, не индуцируя репродукцию, сборку и выход дочерних популяций, такой встроенный вирусный геном называется провирус (рис.20). Встраивание может происходить под действие различных факторов, что приводит к мутациям, злокачественным трансформациям. Подобные ситуации могут возникнуть при гепатите B и C, при ВИЧ-инфекции и гепретических инфекциях (сакрома Капоши).
 |
Рис.20. Интегративная инфекция.
Абортивная инфекция. Абортивной называется инфекция, которая не завершается образованием инфекционных вирусных частиц, или они образуются в гораздо меньшем количестве, чем при продуктивной инфекции (рис.21). Абортивная инфекция может возникнуть при следующих трех обстоятельствах: 1) заражение чувствительных клеток дефектным вирусом; 2) заражение чувствительных клеток в неразрешающих условиях; 3) заражение нечувствительных клеток стандартным вирусом.

Рис.21. Абортивная инфекция.
Заражение чувствительных клеток дефектным вирусом. Дефектным называется такой вирус, который не способен проявить все генетические функции, необходимые для образования инфекционного потомства.
Существуют дефектные вирусы и дефектные вирусные частицы. Дефектными называются такие вирусы, которые репродуцируются лишь в присутствии вируса-помощника, например аденоассоциированный вирус (семейство парвовирусов), дающий потомство только в присутствии аденовируса-помощника. Дефектные вирусные частицы накапливаются в популяции многих вирусов, особенно при пассировании их с высокой множественностью инфекции. Дефектные частицы интерферируют при репродукции вируса с инфекционными вирусными частицами и потому называются дефектными интерферирующими частицами (ДИ-частицами). Этот тип вирусных частиц наиболее хорошо изучен на модели вирусов везикулярного стоматита и гриппа. Получение дефектных частиц вируса гриппа при заражении куриных эмбрионов с высокой множественностью инфекции получило название феномена фон Магнуса по имени исследователя, впервые его описавшего. Дефектные вирусные частицы вызывают абортивную инфекцию в связи с тем, что они лишены части генетического материала. Например, дефектные частицы вируса гриппа содержат неполные последовательности Р-генов, кодирующих три высокомолекулярных вирусных белка.
Заражение чувствительных клеток в неразрешающих условиях. Абортивная инфекция может возникать при изменении условий, в которых происходит инфекционный процесс. Эти условия возникают в организме и могут моделироваться в эксперименте; в организме — повышение температуры, изменение рН в очаге воспаления и концентрации ионов, наличие антиметаболитов, ингибиторов и т. д.; в эксперименте — изменение температуры инкубации, состава питательной среды, внесение антиметаболитов и ингибиторов и т. д. В результате клетка либо погибнет без продукции инфекционного вируса, либо инфекция прерывается на определенном этапе. При устранении неразрешающих условий абортивная инфекция превращается в продуктивную. Смена абортивной инфекции на продуктивную может осуществиться и с помощью вируса-помощника.
Заражение нечувствительных клеток стандартным вирусом приводит к наиболее распространенной форме абортивной инфекции. Непермиссивность клетки к определенному вирусному агенту может проявиться на любом этапе инфекции. Чувствительность клетки к ряду вирусов определяется наличием на клеточной поверхности специфических рецепторов, обусловливающих адсорбцию и проникновение вируса в клетку. Такой генетически обусловленный механизм клеточной резистентности наиболее четко установлен для пикорнавирусов, а также онковирусов птиц. Для большинства вирусов можно подобрать две клеточные системы, в одной из которых будет развиваться продуктивная, а в другой — абортивная инфекция. Механизм генетически обусловленной резистентности клеток к вирусам широко варьирует, но в основе его лежит либо отсутствие клеточных факторов, необходимых для репродукции вируса, либо наличие факторов, нарушающих процесс репродукции.
У сложно устроенных вирусов клеточная непермиссивность часто проявляется на стадии сборки вирусных частиц; нарушение сборки в некоторых непермиссивных системах для вирусов гриппа и парамиксовирусов обусловлено уменьшением количества молекул матриксного белка вируса.
Острая и хроническая инфекция. Как продуктивная, так и абортивная инфекция может протекать в виде острой или хронической инфекции.
Острой называется такая форма инфекции, при которой после образования вирусного потомства клетка либо погибает, либо выздоравливает и не содержит вирусных компонентов. Хроническая инфекция — это такая форма инфекции, при которой клетка продолжает продуцировать вирусные частицы или вирусные компоненты в течение длительного времени и передает эту способность дочерним клеткам.
Чаще хроническую форму приобретает абортивная инфекция, так как вирусный генетический материал обычно не входит в состав вирусного потомства, а накапливается в клетках и передается в дочерние клетки. Одним из факторов, вызывающих хроническую инфекцию, являются ДИ-частицы. Такие частицы, попадая в клетки вместе с инфекционными вирусными частицами, конкурируют с ними за факторы репродукции и препятствуют образованию инфекционного потомства. В результате гибель клеток предотвращается. При появлении в системе новых чувствительных клеток в них вновь возникает продуктивная инфекция с образованием ДИ-частиц, и такой цикл инфекции возобновляется снова и снова.
Цитолитическая и нецитолитическая инфекции. Острая инфекция на клеточном уровне может быть цитолитической и нецитолитической в зависимости от судьбы зараженной клетки. Инфекция, завершающаяся гибелью (лизисом) клетки называется цитолитической. Инфекция, которая непосредственно не приводит к лизису клетки, и клетка еще может функционировать в течение некоторого периода времени, продуцируя вирусные частицы, называется нецитолитической.
Смешанная инфекция. В естественных условиях распространен феномен смешанной инфекции, при котором клетка заражается двумя или несколькими разными вирусами. Два и больше инфекционных процесса, происходящих одновременно в одной клетке, могут оказывать различное влияние друг на друга. Возможны несколько вариантов взаимодействия вирусов в процессе смешанной инфекции.
1. Один из вирусов подавляет репродукцию второго вируса, или подавляется репродукция обоих вирусов. Этот феномен называется интерференцией вирусов.
2. Вирус усиливает репродукцию второго вируса в результате комплементации или экзальтации. Комплементация может происходить между двумя родственными или неродственными вирусами, например между аденовирусом и аденоассоциированным вирусом человека или SV40, при этом вирус-помощник предоставляет другому вирусу неструктурный белок. Экзальтация может быть связана с подавлением процесса образования интерферона первым вирусом.
3. Оба вируса не оказывают существенного влияния на процесс репродукции каждого из них, однако может происходить нарушение морфогенеза обоих вирусов.
В основу классификации положены четыре фактора:
1) генерализация вируса;
2) продолжительность инфекции;
3) проявление клинических симптомов;
4) выделение вируса в окружающую среду.
Основанная на этих признаках классификация инфекций, как и любая другая, в известной мере условна, поскольку одна форма может перейти в другую, например, очаговая инфекция — в генерализованную, острая инфекция — в хроническую, латентная — в хроническую и т. д.
Очаговая и генерализованная инфекции. Вирусные инфекции можно разделить на две большие группы: 1) очаговые, когда действие вируса проявляется у входных ворот инфекции в связи с его локальной репродукцией; 2) генерализованные, при которых после ограниченного периода репродукции вируса в первичных очагах происходит генерализация инфекции, и вирус достигает чувствительных тканей, формируя вторичные очаги инфекции.
Очаговые инфекции имеют более короткий инкубационный период, чем генерализованные, защитными факторами организма при этих инфекциях являются скорее секреторные антитела класса IgA, чем антитела гуморальные, а эффективными вакцинами — те, которые стимулируют образование секреторных антител. При генерализованных инфекциях большее значение в защите организма имеют гуморальные антитела. Примером очаговых инфекций являются респираторные и кишечные вирусные инфекции, примером генерализованных — оспа, корь, полиомиелит. Сравнительная характеристика очаговых и генерализованных инфекций представлена в табл. 8. Примером генерализованной инфекции является корь, а очаговой — заболевания, вызываемые респираторно-синцитиальным вирусом, и другие острые респираторные вирусные инфекции.
| Свойства инфекции | Очаговые инфекции | Генерализованные инфекции |
| Место патологического процесса | Входные ворота | Системы органов и тканей |
| Инкубационный период | Относительно короткий | Относительно длинный |
| Наличие вирусемии | Редко | Обычно |
| Продолжительность иммунитета | Кратковременный или неизученный | Обычно длительный |
| Иммунные механизмы | Секреторные антитела (IgA). Локальный клеточный иммунитет | Гуморальные антитела (IgG, IgM), системный клеточный иммунитет |
Острая и персистентная инфекции. Острая инфекция длится относительно непродолжительный период времени и протекает с выделением вирусов в окружающую среду. Окончание инфекции сопровождается элиминацией вирусов благодаря иммунным механизмам. Инфекция может протекать как в клинической, так и в инаппарантной форме. Острая инфекция может завершиться выздоровлением или гибелью организма. Она соответствует продуктивной инфекции на уровне клетки. При продолжительном взаимодействии вируса с организмом возникает персистентная форма инфекции (от лат. persistentia — упорство, постоянство).
Один и тот же вирус может вызвать как острую, так и персистентную инфекцию в зависимости от состояния организма и в первую очередь его иммунной системы. Например, вирус кори может вызвать как острую инфекцию, так и медленную (длительно текущую) — подострый склерозирующий панэнцефалит. Вирусы герпеса, гепатита В и аденовирусы могут вызвать острую и персистентную инфекции и т. д.
Персистентные инфекции могут быть латентными, хроническими или медленными в зависимости от выделения вируса в среду и проявления симптомов заболевания.
Латентная инфекция — это скрытая инфекция, не сопровождающаяся выделением вирусов в окружающую среду. При латентных инфекциях вирус не всегда удается обнаружить либо в связи с его дефектным состоянием, либо в связи с персистенцией субвирусных компонентов, либо в связи с интеграцией клеточным геномом. При воздействии ряда активирующих инфекцию факторов может произойти активация вируса, и латентная инфекция может перейти в острую или хроническую. Латентные инфекции могут вызывать аденовирусы, вирусы герпеса, онкогенные вирусы, вирус СПИД и др.
Хронической инфекцией называется длительно текущий патологический процесс, характеризующийся периодами ремиссий, перемежающимися с периодами обострения, когда вирус выделяется в окружающую среду. Примерами хронической инфекции являются герпетическая, аденовирусная инфекции, хроническая форма вирусных гепатитов и т. д.
Медленные инфекции — это своеобразное взаимодействие определенных вирусов с организмом, характеризующееся длительным инкубационным периодом, тянущимся многие месяцы и даже годы, и последующим медленным, но неуклонным развитием симптомов заболевания, ведущим к тяжелому нарушению функций органов и летальному исходу. К медленным инфекциям относятся медленно прогрессирующие заболевания, в частности, заболеванияЦНС со спонгиоформными энцефалопатиями у человека — куру, болезнь Крейтцфельдта-Якоба (пресенильная деменция), а у животных — трансмиссивная энцефалопатия норок и скрепи у овец.
К медленным инфекциям относят также подострый склерозирующий панэнцефалит, который вызывается вирусом кори, рассеянный склероз, амиотрофический боковой склероз и некоторые другие заболевания человека и животных.
При некоторых медленных инфекциях существенную роль играют генетические механизмы (скрепи, куру, амиотрофический боковой склероз), при других — иммунопатологические механизмы (подострый склерозирующий панэнцефалит, алеутская болезнь норок, лимфоцитарный хориоменингит).
Персистентные инфекции являются серьезной проблемой современной вирусологии и медицины. Большинство вирусов человека и животных способны персистировать в организме и вызывать латентные и хронические инфекции, и удельный вес персистентных инфекций намного превышает таковой острых инфекций. При персистентных инфекциях постоянно или периодически происходит выделение вирусов в окружающую среду, и персистентные инфекции являются основным фактором эпидемизации населения. Персистенция вирусов обусловливает их сохранение как биологического вида и является причиной изменчивости свойств вирусов и их эволюции.
Большую роль персистенция вирусов играет в перинатальной патологии. Вертикальная передача персистирующего вируса от инфицированной матери плоду и активная репродукция вируса в его тканях особенно опасны в первые месяцы беременности, так как приводят к аномалиям развития плода или его гибели. К числу таких вирусов относятся вирусы краснухи, простого герпеса, ветряной оспы, цитомегалии, Коксаки В и ряд других.
Борьба с персистентными инфекциями затруднена из-за отсутствия адекватных подходов к их лечению и профилактике.

