 2014-02-05
2014-02-05 4067
4067Общие положения
Опухоли способны возникать спонтанно, без видимых на то причин, а также могут быть индуцированы. У человека, как правило, появляющиеся опухоли – это спонтанные, хотя отличить спонтанно возникшую опухоль от индуцированной чрезвычайно трудно. Опухоли, искусственно вызванные, называются индуцированными. Индуцировать канцерогенез способны различные химические вещества (метилхолантрен, бензпирен, афлатоксин В1), физические факторы (ультрафиолетовый свет, ионизирующая радиация), вирусы (папова-, герпес-, адено- и ретровирусы). У человека 3 типа вирусов тесно связаны с опухолями, это вирус Эпштейна-Барр (лимфома Беркита), цитомегаловирус (саркома Капоши) и вирус папилломы.
В основе злокачественной трансформации клеток лежат изменения в их геноме. К озлокачествлению клеток могут приводить мутации, изменяющие генетический материал, либо реактивация «молчащих» эмбриональных генов, усиление или подавление функционирования «нормальных» генов, ответственных за пролиферацию и дифференцировку клеток, а также появление новых генов, как это наблюдается при вирусной индукции канцерогенеза. Гены, связанные с опухолевой трансформацией клеток, получили название онкогенов; в нормальных клетках они представлены в виде проонкогенов. Проонкогены в нормальных клетках функционируют как важные регуляторы их жизнедеятельности. Они кодируют молекулы, контролирующие и регулирующие клеточный рост, дифференцировку клеток, процессы проведения регуляторных (активационных и супрессирующих) сигналов внутрь клетки, процессы трансляции, нуклеинового обмена, апоптоза.
Известные к настоящему времени проонкогены и кодируемые ими молекулы приведены в таблице 6-1. По своим функциональным особенностям они могут быть разделены на три категории: 1) гены, индуцирующие клеточную пролиферацию; 2) гены, подавляющие клеточную пролиферацию, опухолесупрессирующие гены; 3) гены апоптоза.
Таблица 6-1. Некоторые проонкогены и кодируемые ими молекулы.
| Гены | Кодируемые продукты |
| Гены, кодирующие факторы, способствующие клеточной пролиферации | |
| 1. Ростовые факторы sis 2. Рецепторы факторов роста fms erbB neu erbA 3. Трансдукторы srs abl Ha-ras N-ras K-ras 4. Факторы транскрипции myc jun fos | тромбоцитарный ростовой фактор (PDGF) рецептор колониестимулирующего фактора 1 (CSF-1) рецептор эпидермального фактора роста (EGF) белок (HER2), связанный с EGF- рецептором рецептор тиреоидного гормона тирозинкиназа тирозинкиназа ГТФ-связанные белки с ГТФ-азной активностью ГТФ-G-связанные белки с ГТФ-азной активностью ГТФ-связанные белки с ГТФ-азной активностью ДНК-связанные белки компонент фактора транскрипции АР1 компонент фактора транскрипции АР1 |
| Антиопухолевые гены (гены, кодирующие продукты, подавляющие клеточную пролиферацию) | |
| Rb p53 DCC APC NF1 | ингибитор ретинобластомы ядерный белок, подавляющий образование мелкоклеточного рака легких и толстой кишки супрессор карциномы толстой кишки супрессор аденоматозного полипоза супрессор нейрофиброматоза |
| Гены, контролирующие апоптоз | |
| bcl-2 | супрессор апоптоза |
Первая категория проонкогенов и их онкогенных копий кодирует белки, которые функционируют как факторы роста (тромбоцитарный фактор роста (PDGF) или рецепторы к факторам роста (рецептор колониестимулирующего фактора 1 (R-СSF1)), рецептор эпидермального фактора роста (R-EGF), рецептор тиреоидного гормона). Обычно одна популяция клеток продуцирует ростовые факторы, а другая экспрессирует рецепторы к ним. В нормальных условиях эти процессы тщательно регулируются и тонко сбалансированы. Установлено, что повышенная продукция факторов роста или повышенная экспрессия рецепторов, их воспринимающих, способны приводить к повышенному и неконтролируемому клеточному росту.
Другие продукты, кодируемые этой категорией проонкогенов, участвуют в трансдукции сигнала с поверхности внутрь клетки (тирозинкиназы, ГТФ-связанные белки с ГТФ-азной активностью) и в процессах транскрипции (ДНК-связанные белки). Повышенная активность каждого из названных проонкогенов способна стимулировать клеточный рост.
Вторая категория проонкогенов – антионкогены кодируют белки, подавляющие клеточный рост. Инактивация этих генов приводит к развитию опухоли. К ним относятся гены Rb, p53, DCC, APC, NF1. Например, известно, что инактивация гена Rb приводит к развитию у детей редкого наследственного заболевания, такого, как ретинобластома. Опухоль развивается из предшественников нервных клеток сетчатки. Мутация в гене р53, который кодирует ядерный фосфопротеин, приводит к развитию в 90% случаев мелкоклеточного рака легкого и более чем в 50% случаев рака груди и рака толстой кишки.
Третья категория проонкогенов кодирует белки, регулирующие запрограммированную клеточную смерть (подавляющие или индуцирующие апоптоз). В этой категории генов хорошо изучен ген bcl-2. Установлено, что bcl-2 является важным регулятором продолжительности жизни гемопоэтических клеток, Т- и В-лимфоцитов. Интересно, что вирус Эпштейна-Барр также содержит ген, подобный bcl-2, и через инкорпорацию этого гена способен вызывать супрессию апоптоза в инфицированных клетках и тем самым нарушать баланс между процессами естественной гибели клеток и их новообразованием.
Также онкогены были выявлены у ряда вирусов. Так, у вируса саркомы Роус был идентифицирован онкоген, названный v-src, который кодирует протеинкиназу 60-кD, которая катализирует присоединение фосфата к остаткам тирозина на белках. При переносе этого онкогена в нормальные клетки последние подвергаются злокачественной трансформации.
Серией работ показано, что превращение нормальных клеток в опухолевые может происходить в результате хромосомной транслокации. Такой пример приведен на рисунке 6-1.
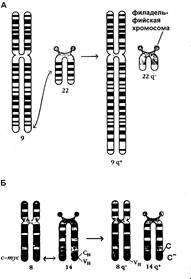
Рис.6-1. Хромосомные транслокации при хроническом миелолейкозе (А) и лимфоме Беркита (Б)
Как видно на рисунке, при лимфоме Беркита ген c-myc (клеточный проонкоген) перемещается со своего нормального положения в хромосоме 8 в кластер гена тяжелой цепи иммуноглобулина на хромосому 14. В результате этой транслокации происходит активация этого гена и усиление синтеза кодируемого им с-белка, который, как известно, функционирует как фактор транскрипции. При хронической миелоидной лейкемии клетки больных содержат филадельфийскую хромосому, которая образуется в результате транслокации генов между 9 и 22 хромосомами.
Суммируя накопленный к настоящему времени материал, можно заключить, что трансформация нормальных клеток в опухолевые может происходить в результате встраивания вирусных онкогенов в геном клетки, хромосомных транслокаций проонкогенов, что приводит к их активации и трансформации клеточных проонкогенов в онкогены под влиянием химических канцерогенов, физических факторов, вирусной инфекции. Все эти процессы сопровождаются нарушением нормального функционирования генов, которые в нормальных условиях выполняют важные регуляторные функции. В результате изменения функциональной активности генов происходит изменение количества (увеличение или снижение) регуляторных молекул и их активности, что в итоге приводит к расбалансировке между процессами размножения клеток и их гибелью.
Приведенное заключение подтверждается тем, что во многих опухолевых клетках, в отличие от нормальных, содержатся множественные копии клеточных онкогенов, ответственных за клеточную пролиферацию, что в свою очередь способно приводить к увеличению продукции молекул, усиливающих клеточный рост. Так, во многих раковых клетках выявляется повышенная экспрессия рецепторов к эпидермальному фактору роста, который кодируется геном c-erbB, а при раке груди наблюдается усиленный синтез рецепторов к фактору роста, кодируемого c-neu.

