 2014-02-03
2014-02-03 4034
4034Лекция-6
План: ПОЛИГИБРИДНОЕ СКРЕЩИВАНИЕ
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Установив закономерности наследования признаков при моногибридном скрещивании, Мендель приступил к проведению ди-гибридного скрещивания. Он отобрал два сорта гороха, которые отличались по двум парам альтернативных признаков. Одна из них определяла форму семян: круглая (АА) или морщинистая (аа), другая — окраску: желтая (ВВ) или зеленая (bb). При опылении растений с круглыми желтыми семенами (ААВВ) пыльцой сорта с морщинистыми зелеными семенами (aabb) все семена гибридов первого поколения оказались круглыми и желтыми (АаВЬ):
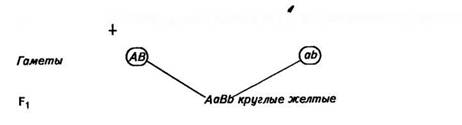
р О ААВВ круглые желтые х О aabb морщинистые зеленые
Доминировали та же форма и тот же цвет семян, что и при моногибридном скрещивании. При самоопылении 15 гибридных растений из Fi с круглыми желтыми семенами (АаВЬ х АаВЬ) во втором поколении было получено 556 семян, которые по парам признаков распределились в следующем количественном соотношении: круглых желтых —315, круглых зеленых— 108, морщинистых желтых — 101, морщинистых зелены*—32. Мендель Принял число 32 за единицу и получил формулу расщепления по фенотипу в F2 при дигибридном скрещивании — 9 круглых желтых: 3 круглых зеленых: 3 морщинистых желтых: 1 морщинистую зеленую. Подсчет отдельно по каждой паре признаков показал, что по форме 423 растения были с круглыми, 133 с морщинистыми семенами, отношение, близкое 3:1; по окраске 416 растений было с желтыми и 140 с зелеными семенами, отношение также 3:1.
Расщепление по каждой паре признаков шло так же, как и при моногибридном скрещивании. Мендель делает вывод о том, что при дигибридном скрещивании в F2 наблюдается сочетание двух моногибридных расщеплений. При сочетании двух моногибридных расщеплений у Fi (AaBb) должно происходить независимое^, распределение в половые клетки аллельных генов по одному из каждой пары. Это приведет к образованию четырех типов гамет (ЛВ, Ab, аВ, ab) в равном числе.
Ниже приводится решетка, предложенная Р. Пеннетом, где вверху по горизонтали и слева по вертикали помещены гаметы родителей, а в центральной части — генотипы потомков F2-
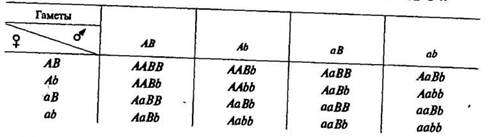
Расщепление гибридов F2 по генотипу дает 9 классов: IAABR2 AaBB-.2AABk4AaBb:lAAbb:2Aabb:laaBB:2aaBb:laabb. По фенотипу образуется четыре класса в отношении: 9 частей круглых желтых (\AABB, lAaBB, lAABb, ААаВЬ), три части круглых морщинистых (lAAbb, 2Aabb), три части морщинистых желтых (ХааВВ, 2ааВЬ), одна часть морщинистых зеленых (aabb).
Анализ по фенотипу показывает, что по форме семян на 12 частей круглых наблюдается 4 части морщинистых (3:1), по окраске на 12 частей желтых —4 зеленых (3:1). Следовательно, во втором поколении дигибридного скрещивания наследование по одной паре признаков (форме семян) идет независимо от наследования по другой паре (окраске семян).
У гибридов F2 наблюдается сочетание признаков во всех возможных комбинациях. Появляются потомки, имеющие признаки обеих родительских форм, например растения с желтыми морщинистыми и зелеными гладкими семенами.
Мендель проверил путем самоопыления генотип всех растений F2 и подтвердил, что расщепление по генотипу при дигибридном скрещивании является результатом независимого комбинирования двух отдельных расщеплений по генотипу при моногибридном скрещивании. Получение формулы расщепления показано в таблице 2, где вверху по горизонтали дано расщепление по генотипу, определяющее форму семян, а по вертикали — определяющее окраску семян.
2. Вывод формулы расщепления по генотипу при дигибридном
скрещивании
| Расщепление | По одной паре аллелей | |||
| по генотипу | АА | 2Аа | аа | |
| По другой паре | ||||
| аллелей | ||||
| ВВ | 1ААВВ | 14а В В | laaBB | |
| 2ВЬ | 2ЛЛВЬ | 4АаВЬ | 2ааВЪ | |
| ЬЬ | lAAbb | 2Aabb | laabb |
Примечание. В центральной части таблицы показано расщепление по генотипу одновременно по двум парам признаков. Оно имеет такой вид: ХААВВ + + lAABb + lAAbb + lAaBB + \AaBb + lAabb + laaBB + laaBb + laabb.
Мендель показал, что независимое наследование признаков обусловлено независимым наследованием разных пар аллелей. В основе независимого наследования разных пар аллелей (наследственных задатков) лежит независимое комбинирование хромосом при образовании гамет.
Правильность своих выводов о независимом комбинировании генов и признаков Мендель подтвердил путем проведения анализирующего скрещивания. Он скрестил гибридные растения Fi с круглыми желтыми семенами (АаВЬ) с отцовским растением, гомозиготным по рецессивным признакам морщинистой формы семян и зеленой их окраски (aabb). У гибрида Fi при независимом комбинировании генов равновероятно образование четырех сортов гамет (АВ, Ab, aB, ab), у отцовского растения образуются гаметы только одного сорта (ab). Наследование идет по следующей схеме:
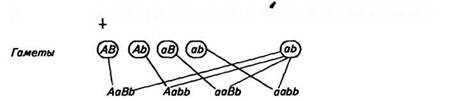
р О АаВЬ круглые желтые х О aabb морщинистые зеленые
Было получено потомство четырех фенотипов: круглые желтые (АаВЬ), круглые зеленые (Aabb), морщинистые желтые (ааВЬ), морщинистые зеленые (aabb). Потомков 'каждого типа было получено одинаковое число — по 25 %. Так как у отцовского растения половые клетки были одинаковые — ab, то равное число особей с каждым фенотипом является результатом того, что гибриды Fi (АаВЬ) образовали половые клетки каждого типа (AB, Ab, аВ, ab) в равном количестве. Это возможно только при независимом комбинировании генов.
На основании опытов по дигибридному скрещиванию был установлен закон независимого наследования признаков (называют также — независимого наследования аллелей разных генов).
Закон независимого наследования признаков состоит в том, что во втором поколении каждая пара аллельных генов и признаков, определяемых ими, ведет себя независимо от других пар аллельных генов и признаков. При этом возникают всевозможные сочетания в определенных числовых соотношениях по фенотипу и генотипу. При дигибридном скрещивании, при полном доминировании, соотношение по фенотипу будет 9:3:3:1, при тригибридном скрещивании будет свое определенное соотношение и т. д.
Разберем пример дигибридного скрещивания применительно к животным. Скрестим свиноматку породы ландрас белую с висячими ушами с хряком беркширской породы черным со стоячими ушами. Одна пара признаков характеризует масть (белая или черная), другая пара — форму ущей (висячие или стоячие). Ген доминантного признака белой масти обозначим прописной буквой А, а аллельный ген рецессивной черной масти — строчной буквой а. Ген доминирующих висячих ушей обозначим буквой Д ген стоячих ушей — Ь. Допустим, что аллельные гены по указанным парам признаков находятся в разных парах хромосом' (рис. 10). Оба родителя гомозиготны: мать по доминантным признакам белой масти и висячих ушей (ААВВ), отец по рецессивным признакам черной масти и стоячих ушей (aabb). В период образования половых клеток при мейозе из каждой пары гомологичных хромосом в гамету придет только одна. Поскольку родители гомозиготны, то у каждого из них будут все гаметы одного типа: у свиноматки АВ, у хряка ab.
В первом поколении в соответствии с первым законом Менделя все свиньи по фенотипу будут одинаковыми: белыми, с висячими ушами, по генотипу дигетерозиготны, т. е. гетерозиготны по обеим парам признаков (АаВЬ).
В гибридах F2, которые получают путем спаривания животных Fi между собой, наблюдается расщепление. Предположим, что одна пара аллельных генов А и а находится в более длинных гомологичных хромосомах, другая пара В и Ъ — в более коротких. В результате мейоза из каждой пары гомологичных хромосом в каждую половую клетку пойдет только одна. Аллели Аи а
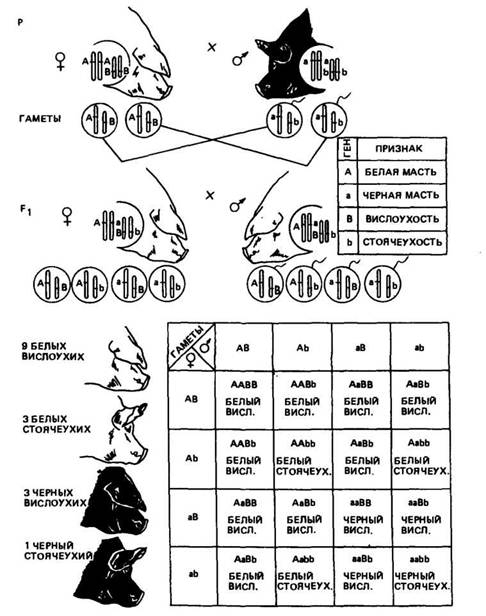
Рис. 10. Схема скрещивания белой с висячими ушами сяиноматки с черным
со стоячими ушами хряком
разойдутся, то же самое произойдет и с аллелями В и Ъ. Расхождение в гаметы хромосом из каждой пары происходит независимо, поэтому аллель А может уйти с равной вероятностью как в те гаметы, куда ушел аллель /?, так и в те, куда ушел аллель Ь. Аллель а также с равной вероятностью может попасть в гаметы как с аллелем В, так и с аллелем Ь. В результате как хряки, так и свиноматки из F[ образуют по четыре сорта гамет: АВ, АЪ, аВ, аЪ в равном количестве.
Каждый из спермиев может оплодотворить любую из яйцеклеток с одинаковой вероятностью. Получается 16 возможных сочетаний гамет отца и матери. Результаты этой случайной встречи гамет хорошо видны при использовании решетки Пен-нета. В верхней горизонтальной строке как заголовки записаны типы гамет одного родителя, а слева вертикально, как заголовки строчек, расположены типы гамет другого родителя. В каждый квадрат на пересечении столбца и строчки записаны генотип и фенотип потомка, определяемые исходя из сочетаний гамет, стоящих в заголовках. В заголовках располагают сначала гаметы с доминирующими генами, затем с доминирующим и рецессивным и в конце с рецессивными. При записи генотипа каждого потомка сначала располагают гены одной аллельной пары, затем другой, рядом указывают фенотип. Затем подсчитывают особей с разными фенотипами и генотипами.
Из данных решетки видно, что в F2 среди особей с разным фенотипом наблюдается следующее количественное соотношение: 9 частей белых с висячими ушами; 3 части белых со стоячими ушами; 3 части черных с висячими ушами; 1 часть черных со стоячими ушами. Рассматривая каждую пару признаков отдельно, находим, что на 12 белых свиней приходится 4 черные (отношение 3:1) и, с другой стороны, на 12 свиней с висячими ушами — 4 со стоячими (отношение 3:1).
Таким образом, каждая пара признаков при наследовании ведет себя независимо от другой пары, и только в результате их свободного комбинирования наблюдается характерное для ди-гибридного расщепления соотношение фенотипов в F2 — 9:3:3:1, которое можно рассматривать как результат сочетания двух моногибридных расщеплений (3:1 и 3:1).
Расщепление по генотипу во втором поколении точно такое же, как установил Мендель на растениях гороха.








