 2014-02-03
2014-02-03 2469
2469СВЯЗАННЫЕ С ХАРАКТЕРОМ ДОМИНИРОВАНИЯ
Типы доминирования. Вскоре после переоткрытия законов Менделя на животных и растениях разных видов было установлено, что не все признаки проявляют полное доминирование. Были выявлены случаи промежуточного наследования, неполного доминирования, сверхдоминирования и кодоминирования.
При промежуточном наследовании потомство в первом поколении сохраняет единообразие, но оно не похоже полностью ни на одного из родителей, как это было при полном доминировании, а обладает признаком промежуточного характера. Например, известно, что среди овец наряду с нормальноухи-ми имеются и безухие. Скрещивание безухих овец (аа) с нор-мальноухими (АА), имеющими длину уха около 10 см, дает в первом поколении потомство (Аа) исключительно с короткими ушами -*- около 5 см.
Иногда признак принимает не среднее выражение, а уклоняется в сторону родителя с доминирующим признаком, тогда говорят о неполном доминировании. Например, при скрещивании коров с белыми пятнами на туловище, белыми брюхом иконечностями с быками со сплошной окраской получается потомство со сплошной окраской, но с небольшими пятнами на ногах или других частях туловища.
При сверхдоминированииу гибридов первого поколения проявляется гетерозис — явление превосходства потомства над родительскими формами по жизнеспособности, энергии роста, плодовитости и продуктивности. Сверхдоминированием в определенной мере объясняется эффект гетерозиса, наблюдаемый при получении в птицеводстве трех- и четырехлинейных гибридов.
При кодоминировании у гибридной особи в равной мере проявляются оба родительских признака. По типу кодоминирования наследуется большинство антигенных факторов довольно многочисленных систем групп крови у домашних животных разных видов и человека. Так же наследуются разные типы белков и ферментов: гемоглобин, амилаза и т. д.
Расщепление по фенотипу 3:1 во втором поколении моногибридного скрещивания наблюдается при полном доминировании признака.
При промежуточном наследовании, неполном доминировании и кодоминировании в результате разного характера взаимодействия аллельных генов гибриды первого поколения (Аа) отличаются по фенотипу от родителя с доминантным признаком (АА). Отсюда и в потомстве F2 гетерозиготные особи будут иметь свойственный для них фенотип. В результате расщепление по фенотипу и генотипу будет одинаковым: 1:2:1. Так, при скрещивании длинноухих и безухих овец в Fi все потомки появляются короткоухими (рис. 9). При скрещивании их между собой (Аа х Аа) во втором поколении одна часть потомков (АА) будет иметь уши длинные, две части (Аа) — короткие и одна часть (аа) рождается безухими. Таким образом, на расщепление по фенотипу во втором поколении влияет характер доминирования признака.
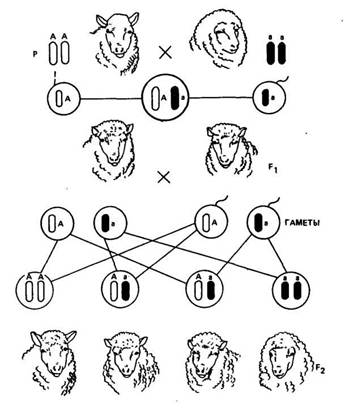
Рис. 9. Схема наследования безухости у овец:
А — ген длинных ушей; а — ген безухости
Летальные гены. Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот F2- Разная жизнеспособность зигот может быть обусловлена наличием ле-tsuibHbix генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими половой зрелости, называются летальными, более 50 % — сублетальными (полулетальными) и менее 50 % — субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2—3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32—35 "С. Но если температура в брудерах будет повышена на 5,5 "С, то погибнет значительно меньше голых цыплят. В 4—5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (YY), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
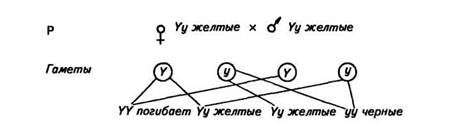
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2—4-й день после рождения от уродства прямой кишки — отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина — гетерозиготной (LI), несущей рецессивный задаток (/) летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (It), с уродством прямой кишки. Они все погибли. (Более подробно об аномалиях при летальных генах будет сказано в соответствующей главе.)

