 2014-02-03
2014-02-03 1929
1929Представление о том, что генетическая информация о структуре белковых молекул зашифрована в ДНК путем определенного расположения нуклеотидов, конкретизировал Ф. Крик в гипотезе последовательности, согласно которой последовательность элементов гена определяет последовательность аминокислотных остатков в полипептидной цепи. Было установлено, что наследственную информацию с ДНК считывает иРНК, которая образуется комплементарно одной из цепей ДНК. Однако не было известно, каким образом переводится нуклеотидная последовательность иРНК в аминокислотную последовательность полипептидной цепи. Можно было предположить, что генетический код не может состоять из одного или двух нуклеотидов, так как их только четыре и сочетаний из двух (421) может быть только 16, а аминокислот 20. Г. Гамов в 1954 г. впервые высказал мысль о том, что генетический код должен быть триплетным. В этом случае получается (43) 64 сочетания, и их вполне достаточно для кодирования всех аминокислот.
Начало экспериментальному анализу природы генетического кода положили М. Ниренберг и Дж. Маттеи в 1961 г. Они создали простейшие синтетические полимеры типа иРНК. Искусственно полученный полимер, содержащий только уридиновые нуклеотиды, в которых основанием является урацил, вводили в бесклеточную среду, полученную из кишечной палочки. В результате был получен полипептид, состоящий только из фенил-аланина — полифенилаланин. Кодон для фенилаланина был расшифрован как УУУ.
К расшифровке генетического кода активно подключился С. Очоа с сотр. В течение 3—4 лет в лабораториях М. Ниренбер-га и С. Очоа был определен состав большинства кодонов. Однако требовалось определить последовательность нуклеотидов в ко-донах. Это удалось сделать при помощи двух методов. Г. Корана с сотр. разработал метод химического синтеза ДНК-подобных полимеров с заданной последовательностью нуклеотидов, что
позволяло получить РНК также с заранее известной последовательностью нуклеотидов и использовать ее в бесклеточной системе белкового синтеза. Второй метод предложили М. Ниренберг и П. Ледер, исходя из того, что промежуточными продуктами при синтезе белка являются аминокислоты, связанные с тРНК. Убедившись в том, что одного триплета иРНК (трех нуклеотидов) достаточно для связывания с рибосомой и тРНК, ученые использовали тринуклеотидные матрицы с известным чередованием оснований для того, чтобы изучить, какую аминокислоту доставит тРНК.
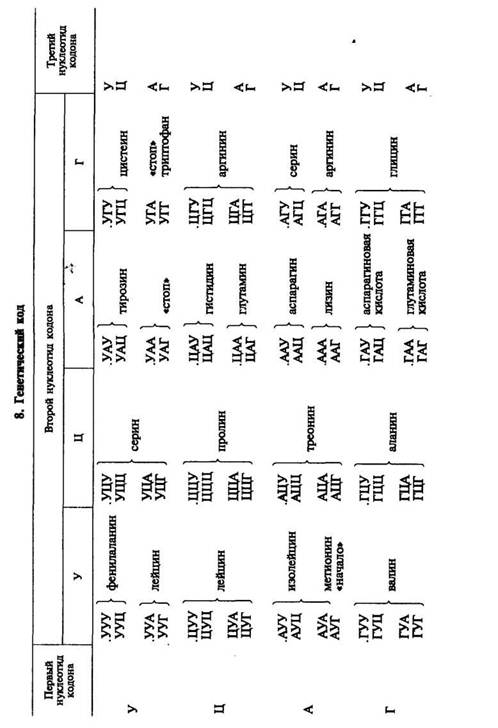
В результате использования методов, разработанных Г. Кора-ной, М. Ниренбергом и П. Ледером, к 196о г. были определены все триплеты, кодирующие ту или иную аминокислоту. Триплет иРНК получил название кодона. Генетический код был полностью расшифрован (табл. 8), значит, была выяснена природа связи между структурой гена и соответствующего белка. Было установлено, что 61 триплет кодирует аминокислоты, 3 триплета не соответствуют никакой аминокислоте и определяют конец трансляции..
Выявлены следующие особенности генетического кода: 1) генетический код триплетный (каждая аминокислота кодируется тремя нуклеотидами); 2) неперекрывающийся (соседние триплеты не имеют общих нуклеотидов); 3) вырожденный (за исключением метионина и триптофана все аминокислоты имеют более одного кодона); 4) универсальный (в основном одинаков для всех живых организмов); 5) в кодонах для одной аминокислоты первые два нуклеотида, как правило, одинаковы, а третий варьирует; 6) имеет линейный порядок считывания и характеризуется колине-арностью, т. е. совпадением порядка расположения кодонов в иРНК с порядком расположения аминокислот в синтезирующейся полипептидной цепи.
Сравнительно недавно выяснилось, что в митохондриях нарушается универсальность генетического кода. Четыре кодона в митохондриях изменили свой смысл: кодон УГА отвечает триптофану, АУА — метионину, а кодоны АГА и АГГ стали терминирующими. В митохондриях синтезируется небольшое количество белков, которые используются ими же. Открытие новых кодонов у митохондрий может служить доказательством того, что код эволюционировал, что он не сразу стал таким, каким мы его знаем теперь.
СИНТЕЗ БЕЛКА В КЛЕТКЕ
В настоящее время можно считать установленным, что наследственность реализуется в процессе биосинтеза белка. Синтез ферментов и других белков, необходимых для жизнедеятельности и развития организмов, происходит в основном на первой стадии
интерфазы, до начала репликации ДНК. В процессе синтеза белка различают этапы транскрипции и трансляции.
Транскрипция заключается в том, что наследственная информация, записанная в ДНК (гене), точно транскрибируется (переписывается) в нуклеотидную последовательность иРНК. Синтез иРНК начинается с участка инициации транскрипции, называемого промотором. Промотор расположен перед геном и включает около 80 нуклеотидов. У вирусов и бактерий этот участок включает около 10 нуклеотидов (один виток спирали). Транскрипция осуществляется с помощью ферментов РНК-полимераз. РНК-полимераза прочно связывается с промотором и €расплавляет» его, разъединяя нуклеотиды комплементарных /цепей. Затем этот фермент начинает двигаться вдоль гена и по мере разъединения цепей ДНК на одной из них, которая являет- ся смысловой, ведет синтез иРНК, согласно принципу компле-ментарности присоединяя аденин к тимину, урацил к аденину, цитозин к гуанину и гуанин к цитозину. Те участки гена, на которых полимераза образовала иРНК, вновь соединяются, а синтезируемая молекула иРНК постепенно отделяется от ДНК. Конец синтеза иРНК определяется участком остановки транскрипции — терминатором. Нуклеотидные последовательности промотора и терминатора узнаются специальными белками, регулирующими активность РНК-полимеразы.
В 1977 г. было обнаружено, что у эукариот в последователь-■ ности нуклеотидов ДНК имеются отрезки, не содержащие информации, которые были названы интронами. Участки ДНК, несущие информацию, называются экзонами.
При считывании информации с определенного участка ДНК (гена) сначала образуется транскрипт всей последовательности (про-мРНК), а затем происходит процесс созревания иРНК, называемый процессингом. При процессинге происходит сплайсинг, который заключается в том, что в ядре интроны из РНК как бы «выпетливаются» и удаляются, а информативные участки — экзо- ны соединяются при помощи ферментов лигаз в одну непрерыв- ную последовательность иРНК. Перед выходом из ядра к началь- ной части иРНК (5'-концу) присоединяется остаток метилиро- ванного гуанина, называемый «колпачком», а к концу иРНК (З'-концу) присоединяется примерно 200 остатков адениловой кислоты. В таком виде зрелая иРНК (матричная РНК) проходит через ядерную мембрану в цитоплазму, где соединяется с рибо- сомой. Считают, что у эукариот «колпачок» иРНК играет роль в связывании с малой субчастицей.
Трансляция заключается в том, что последовательность расположения нуклеотидов в иРНК переводится в строго упоря- доченную последовательность расположения аминокислот в мо- лекуле синтезируемого белка. Процесс трансляции включает два
этапа: активирование аминокислот и непосредственно синтез белковой молекулы.
Активирование свободных аминокислот и присоединение их к тРНК осуществляются при помощи ферментов аминоацил-тРНК-синтетаз. Точность процесса трансляции зависит, по-видимому, в значительной мере от того, с какой точностью каждая синтета-за выберет одну определенную аминокислоту и присоединит ее к соответствующей тРНК. Считается, что в молекуле каждой ами-ноацил-тРНК-синтетазы имеется по крайней мере три дентра связывания: для аминокислоты, тРНК и АТф. Сначала осуществляется связь аминоацил-тРНК-синтетазы с определенной чами-нокислотой, а затем активированная аминокислота присоединяется к акцепторному участку (ЦЦА) транспортной РНК. В результате образуется аминоацил-тРНК (аа-тРНК). Нагруженная аминокислотой тРНК взаимодействует с одним из белковых факторов, который в комплексе с ГТФ необходим для транспорта тРНК к рибосоме и связывания с ней.
В период трансляции происходит реализация генетической информации в процессе синтеза белковой молекулы определенной структуры. Синтез подразделяется на три стадии: инициации, элонгации и терминации.
Инициация. ЕМпериод стадии инициации рибосома сначала представлена двумя отдельными субчастицами, так как для начала процесса необходима рибосома диссоциированная.
Инициация синтеза полипептидной цепи начинается с присоединения малой субчастицы рибосомы к соответствующему центру связывания на иРНК. Сигналом инициации трансляции служит кодон для метиоккна АУТ, который расположен в начале иРНК (рис. 21). К кодону АУТ своим антикодоном УАЦ присоединяется тРНК, нагруженная аминокислотой метионином (у бактерий инициаторной является тРНК, которая переносит фор-милметионин). Затем к комплексу, состоящему из малой субъединицы, иРНК и тРНК, присоединяется большая субъединица рибосомы. В результате образуется полностью собранная рибосома (80S), включающая одну молекулу иРНК и инициаторную тРНК с аминокислотой. В большой субъединице имеется аминоацильный и пептидильный центры. Сначала первая аминокислота (метио-нин) попадает в аминоацильный центр. В процессе присоединения большой субчастицы рибосомы иРНК продвигается на один кодон, тРНК из аминоацильного центра перемещается в пептидильный центр. В аминоацильный центр поступает следующий кодон иРНК, который может принять следующую аминоацил-тРНК. С этого момента начинается вторая стадия трансляции.
Элонгация. В эту стадию многократно повторяется цикл присоединения аминокислот к растущей полипептидной цепи. Так, в аминоацильный центр рибосомы строго в соответствии с кодо-ном иРНК поступает вторая нагруженная тРНК, которая своим

Рис. 21. Схема синтеза, белка в клетке:
вверху — инициация; внизу — элонгация
антикодоном соединяется с комплементарным кодоном иРНК. Сразу же при помощи фермента пептидилтрансферазы предшествующая аминокислота (метионин) своей карбоксильной группой (СООН) соединяется с аминогруппой (NH2) вновь пришедшей аминокислоты. Между ними образуется пептидная связь (—СО— NH—). В результате тРНК, принесшая метионин, освобождается, а в аминоацильном центре к тРНК присоединен уже дипептид. Для дальнейшего процесса элонгации требуется освободить аминоацильный центр. И он освобождается.
В результате процесса транслокации дипептидил-тРНК продвигается из аминоацильного центра в пептидильный. Это происходит благодаря перемещению рибосомы на один кодон при участии фермента транслоказы и белкового фактора элонгации. Освободившаяся тРНК и кодон иРНК, который был связан с ней, выходят из рибосомы. В освободившийся аминоацильный центр следующая тРНК приносит аминокислоту в соответствии с поступившим туда кодоном. Эта аминокислота при помощи пептидной связи соединяется с предыдущей. При этом рибосома снова продвигается еще на один кодон, и процесс повторяется. Полипептидный синтез в рибосоме идет до тех пор, пока в аминоацильный центр не поступит терминирующий кодон.
Терминация. После того как в аминоацильный центр рибосомы поступит терминирующий кодон иРНК (УАА, УАГ или УГА), к нему присоединяется один из белковых факторов терминации и блокируется дальнейшая элонгация цепи. Полипептидная цепь отделяется от тРНК и рибосомы, освобождаются тРНК и иРНК. Рибосомные субъединицы диссоциируют и могут принять участие в синтезе следующей полипептидной цепи.
На одной молекуле иРНК работает не одна рибосома, а многие (до 100). На каждой из рибосом строится полипептидная цепь. У бактерий транскрипция и трансляция связаны между собой и трансляция начинается до завершения синтеза иРНК на ДНК. Образующиеся при синтезе белка полипептидные цепи претерпевают посттрансляционные преобразования и в конечном итоге выполняют специфические функции, принимая участие в определении признаков организма.
Ингибиторы синтеза белка. В последние годы был выявлен целый ряд ингибиторов, вызывающих нарушение реализации генетической информации у микроорганизмов. Примером могут служить антибиотики. Одним из мощных ингибиторов является пуромицин. Он имеет структурное сходство с концевым остатком адей$шовой кислоты в аминоацил-тРНК, легко взаимодействует с А-участком пептидил-тРНК с образованием пептидилпу-ромицина. Пептидилпуромицин нарушает элонгацию, вызывая обрыв реакции. Предполагается, что стрептомицин и неомицин вызывают ошибки в трансляции иРНК, приводящие к нарушению соответствия между кодонами и включаемыми аминокислотами. Например, кодон УУУ вместо фенилаланина начинает кодировать лейцин, в результате образуется аномальный белок, что ведет к гибели бактерий. Тетрациклины являются ингибиторами синтеза белка в 708-рибосоме. Считается, что тетрациклины тормозят связывание аминоацил-тРНК с аминоацильным центром рибосом. Синтез клеточной иРНК тормозит антибиотик рифа-мицин, используемый при лечении туберкулеза. Этот препарат тормозяще действует на ДНК-зависимую РНК-полимеразу путем связывания с ней. Наиболее чувствительна к нему бактериальная РНК-полимераза. Недавно обнаружено и противовирусное действие рифамицина. Его используют при лечении трахомы, которая вызывается ДНК-содержащим вирусом. Известно ингиби-рующее действие на синтез белка у микроорганизмов и целого ряда других антибиотиков.
Контрольные вопросы.
1. В чем состоит биологическая роль нуклеиновых кислот?
2. Как была доказана роль ДНК в наследственности?
3. Как построена ДНК?
4. Каким образом происходит репликация ДНК?
5. Каковы различия молекул иРНК, рРНК, тРНК?
6. Что такое транскрипция?
7. Что означает термин «трансляция»?
8. Как осуществляется синтез полипептида в рибосомах?

