 2014-02-03
2014-02-03 5992
5992Лекция-12
План: СТРОЕНИЕ И ТИПЫ РНК.
ГЕНЕТИЧЕСКИЙ КОД. СИНТЕЗ БЕЛКА В КЛЕТКЕ
Нуклеиновые кислоты впервые открыл И. Ф. Мишер в 1868 г. Он выделил из ядер клеток особое вещество кислотной природы и назвал его нуклеином. Впоследствии ему дали название «нуклеиновая кислота». Было обнаружено два типа нуклеиновых кислот. Их назвали в зависимости от углеводного компонента, входящего в состав. Нуклеиновую кислоту, в состав которой входит углевод дезоксирибоза, назвали дезоксирибонуклеиновой кислотой (ДНК), а в состав которой входит углевод рибоза, — рибонуклеиновой кислотой (РНК)- В период с 1900 по 1932 г. был определен химический состав нуклеиновых кислот. Они включают следующие компоненты:
РНК
Аденин, гуанин Цитозин, урацил Рибоза
ДНК
Аденин, гуанин Цитозин, тимин Дезоксирибоза
Пуриновые основания Пиримидиновые основания Углеводный компонент
Обе нуклеиновые кислоты включают остатки фосфорной кислоты. Различие заключается в том, что в состав РНК входит азотистое основание урацил вместо тимина и рибоза вместо дезоксирибозы.
В 1936 г. на кафедре биохимии растений Московского университета А. Н. Белозерский с И. И. Дубровской впервые выделили ДНК в чистом виде из растительного материала. К середине 40-х годов было выяснено, что ДНК и РНК одновременно присутствуют в каждом живом организме.
В конце 40-х — начале 50-х годов при изучении нуклеиновых кислот стали использовать новые физические и химические методы исследования. В 1950 г. Э. Чаргафф установил правила нукле-отидных отношений, лежащие в основе строения всех ДНК.
Правила Чаргаффа заключаются в том, что в ДНК содержание аденина равно содержанию тимина (А = Т), а содержание гуанина равно содержанию цитозина (Г = Ц), отсюда А + Г/Т + Ц = 1; сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклео-тидов. В соответствии с этим правилом нуклеотидный состав разных организмов может варьировать только по величине А + Т/Г+ Ц.
К 1952 г. Р. Франклин и М. Уилкинс добились получения высококачественных рентгенограмм ДНК, показавших, что она имеет форму спирали и двойственную структуру.
В 1953 г. Дж. Уотсон и Ф. Крик, опираясь на данные рентге-ноструктурного анализа и правила Чаргаффа, установили структуру ДНК. Согласно их модели молекула ДНК имеет двойную спираль, состоящую из двух полинуклеотидных цепей с общей осью (рис. 17). Диаметр двойной спирали ДНК равен 2 нм, а расстояние между витками 3,4 нм. На каждый виток спирали приходится 10 пар нуклеотидов, отсюда расстояние между азотистыми основаниями равно 0,34 нм.
Структурными единицами полинуклеотидных цепей являются нуклеотиды. В состав нуклеотида входят: одно из азотистых оснований — пуриновое (аденин или гуанин) или пиримидиновое (тимин или цитозин), дезоксирибоза, фосфатный остаток. Эти компоненты соединены друг с другом в следующем порядке: азотистое основание — дезоксирибоза — фосфатный остаток. Соединение одного из оснований с дезоксирибозой приводит к образованию нуклеозида. В случае присоединения фосфатной группы к углеводной части нуклеозида образуется нуклеотид.
Дезоксирибоза в нуклеотидах соединяется с основаниями гли-козидной связью, а с фосфорной кислотой — эфирными связями. Следовательно, по химическому составу любой нуклеотид — это фосфорный эфир нуклеозидов. В соответствии с этим нукле-отиды называются дезоксиадениловой, двзоксигуаниловой, дезокси-цитидиловой и тимидиловой кислотами.
Наряду с главными азотистыми основаниями ДНК содержит также метилированные основания, такие, как 5-метилцитозин, 5-оксиметилцитозин и др. У животных количество 5-метилцито-зина в ДНК обычно не превышает 1,5—2 %.
В каждой из цепей ДНК нуклеотиды последовательно соединены друг с другом с помощью остатка фосфорной кислоты и молекулы дезоксирибозы. Дезоксирибоза связывается с одной молекулой фосфорной кислоты через углерод в положении 3', а с другой — через углерод 5', образуя углеводно-фосфатный остов (рис. 18).
Обе цепи в молекуле ДНК имеют противоположную полярность. Это означает, что межнуклеотидная связь в одной цепи имеет направление 5'-*3', а в другой 3'-»5'.
Азотистые основания нуклеотидов обеих цепей ДНК заключены внутри между витками спирали и соединены водородными связями. В соответствии с правилами Чаргаффа аденин одной цепи связан только с тимином другой цепи, а гуанин — только с цитозином. Пара аденин — тимин соединена двумя водородными связями, а пара гуанин — цитозин — тремя. Такой порядок соответствия азотистых оснований (А **Г и Г **Ц) называется
комплементарностью, и, следовательно, цепи в ДНК комплементарны, они взаимно дополняют друг друга.
Углеводно-фосфатный остов по всей длине во всех молекулах ДНК имеет однотипную структуру и не может нести генетической информации. В противоположность этому рас-
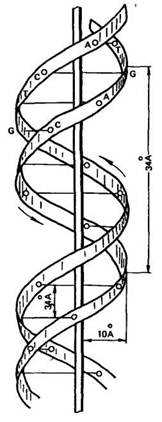
Рис. 17. Схема строения молекулы ДНК из
двух спирально закрученных ценей (по Д. Уотсо-
ну в Ф, Кряку) (Цифры указывают на расстоя-
ним в А между разными точками молекулы)
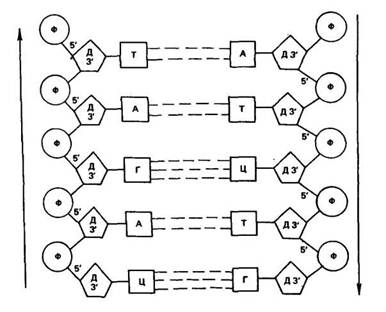
Рис. 18. Схема отрезка двухцепочной молекулы ДНК (по С. М. Гершензону)
положение пуриновых и пиримидиновых оснований нуклеотидов вдоль цепи ДНК очень изменчиво и характерно для каждого данного типа молекул ДНК. Значит, наследственная информация зашифрована различной последовательностью оснований.
Нуклеотидный состав ДНК значительно варьирует в зависимости от принадлежности организма к той или иной систематической группе (табл. 7). Специфичность ДНК выражается соотношением А + Т/Г + Ц, получившим название коэффициента видовой специфичности.
7. Нуклеотидный состав ДНК из различных источников (по А. Ленинджеру, 1976)
| Нуклеотидный | состав, к | ЛОЛ. % | Коэффициент | ||
| Объект | специфичности | ||||
| A | Г | Ц | Т | А + Т/Г + Ц | |
| Человек | 30,9 | 19,9 | 19,8 | 29,4 | 1,52 |
| Животные: | |||||
| овиа | 29,3 | 21,4 | 21,0 | 28,3 | 1,36 |
| курица | 28,8 | 20,5 | 21,5 | 29,2 | 1,38 |
| Растения, грибы, зерна пшеницы | 27,3 | 22,7 | 22,8 | 27,1 | 1,19 |
| Дрожжи | 31,3 | 18,7 | 17,1 | 32,9 | 1,79 |
| Aspergillus niger | 25,0 | 25,1 | 25,0 | 24,9 | 1,00 |
| Бактерии: | |||||
| Е. coli | 24,7 | 26,0 | 25,7 | 23,6 | 0,93 |
| Staphylococcus ayreus | 30,8 | 21,0 | 19,0 | 29,2 | 1,50 |
| Clostridium perfnngens | 36,9 | 14,0 | 12,8 | 36,3 | 2,70 |
| Brucella abortus | 21,0 | 29,0 | 28,9 | 21,1 | 0,72 |
| Sarcina lutea | 13,4 | 37,1 | 37,1 | 12,4 | 0,35 |
В ДНК животных наблюдается избыток А + Т по отношению к Г + Ц. У грибов и бактерий встречаются формы как богатые А + Т, так и с преобладанием Г + Ц, в то же время есть близкие по коэффициенту специфичности к животным. Это говорит о том, что изменчивость в расположении оснований уже достаточна для того, чтобы обеспечить различия между генами этих организмов.
Молекулы ДНК состоят примерно из 2-103—1-Ю8 и более
нуклеотидов и имеют большую относительную молекулярную массу.
Репликация (удвоение) ДНК. ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. Дж. Уотсон и Ф. Крик предложили схему удвоения ДНК, согласно которой спиралевидная двухцепочная ДНК сначала раскручивается (расплетается) вдоль оси. При этом водородные связи между азотистыми основаниями рвутся и цепи расходятся. Одновременно к нуклеотидам каждой цепи пристраиваются комплементарные азотистые основания нуклеотидов второй цепи, где против аденина встает тимин, против тимина — аденин, против гуанина — цитозин и т. д., которые с помощью ферментов ДНК-полимераз связываются в новые полинуклеотидные цепи. В результате из одной образуются две новые дочерние молекулы ДНК. Каждая дочерняя молекула, наследуя структуру одной цепи материнской молекулы, строго сохраняет специфичность заключенной в ней информации. Поскольку матрицей для репликации служит одна из двух цепей молекулы, такой тип синтеза ДНК носит название полуконсервативной ауторепродукции.
Дальнейшие исследования показали, что репликация бактериальных и других молекул ДНК начинается в определенной точке старта. В хромосомах эукариот обнаружено по нескольку таких начальных точек. Цепи ДНК в точке инициации репликации разъединяются под влиянием особого белка геликазы (рис. 19). Возникают одноцепочные участки ДНК, которые становятся матрицами для репликации-притяжения комплементарных нуклеотидов. Эти одноцепочные участки связываются с особыми белками, которые их стабилизируют (препятствуют их комплементарному взаимодействию). Особый фермент топоизомераза (у прокариот назьшается ДНК-гиразой) способствует расщеплению спирали ДНК в области репликационной вилки.
Репликация на материнской цепи, идущей от точки старта в направлении 5'->3', идет в виде сплошной линии. Эта цепь получила название лидирующей. Синтез на второй цепи 3'->5' идет отдельными фрагментами в противоположном направлении (тоже 5'->3')- Эта цепь получила название запаздывающей. Фрагментами являются небольшие участки ДНК (у кишечной палочки около 2000 нуклеотидов, у эукариот около 200). Они называются по
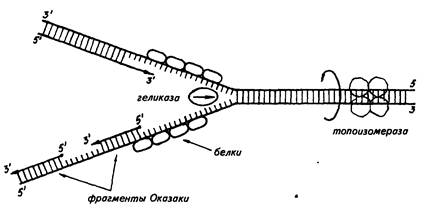
Рис. 19. Синтез лидирующей (вверху) и запаздывающей (внизу) цеией ДНК в вилке репликации (по В. Alberts, R. Sternglanz)
имени открывшего их японского ученого Р. Оказаки. После завершения синтеза фрагменты Оказаки соединяются при помощи фермента ли газы в общую полинуклеотидную цепочку. У эукариот репликация ДНК и соединение различных ее репликационных участков происходят в фазе S-периода интерфазы. После завершения этой фазы в каждой хромосоме имеется две молекулы ДНК, которые становятся двумя идентичными хроматидами.
Структура, способная к репликации (хромосома, плазмида, вирусный геном), называется репликоном.
Самоудвоение молекул ДНК — основа устойчивости генетической информации данного вида и обеспечивает материальную непрерывность наследственного вещества клетки.
СТРОЕНИЕ И ТИПЫ РНК
Многочисленными исследованиями было установлено, что синтез белка в клетке происходит не в ядре, где находится ДНК, а в цитоплазме. Следовательно, сама ДНК не может служить матрицей для синтеза белка. Вставал вопрос о молекулярных механизмах переноса информации, закодированной в ДНК (генах), из ядра в цитоплазму к месту синтеза белка. Сравнительно недавно выяснилось, что молекулами, ответственными за считывание и перенос информации, а также за преобразование этой информации в последовательность аминокислот в структуре белковой молекулы, являются рибонуклеиновые кислоты (РНК). Молекулы рибонуклеиновой кислоты имеют одну полинуклеотидную цепь, Нуклеотиды молекулы РНК называются адениловой гуаниловой, уридиловой и цитцдиловой кислотами. На долю РНК приходится около 5—10 % общей массы клетки.
Существует три основных вида РНК: информационная (иРНК), или матричная (мРНК), рибосомная (рРНК) и транспортная (тРНК). Они различаются по величине молекул и функциям. Все типы РНК синтезируются на ДНК при участии ферментов — РНК-полимераз. Информационная, или матричная, РНК составляет 2—3 % всей клеточной РНК, рибосомная — 80—85, транспортная — около 15 %.
Информационная РНК (иРНК) впервые была обнаружена в 1957 г. Роль ее в том, что она считывает наследственную информацию с участка ДНК (гена) и в форме скопированной последовательности азотистых оснований переносит ее в рибосомы, где происходит синтез определенного белка. Каждая из молекул иРНК по порядку расположения нуклеотидов и по размеру соответствует гену в ДНК, с которого она была транскрибирована. В среднем иРНК содержит 1500 нуклеотидов (75— 3000). Каждый триплет (три нуклеотида) на иРНК называется кодоноц. От кодона зависит, какая аминокислота встанет в данном ме*сте при синтезе белка.' Информационная РНК может обладать относительной молекулярной массой от 250 до 1000 тыс. Д (дальтон).
Существует большое разнообразие иРНК как в отношении состава, так и величины молекулы. Это связано с тем, что в клетке находится большое количество разнообразных белков, а строение каждого белка обусловлено своим геном, с которого иРНК считала информацию.
Транспортная РНК (тРНК) обладает относительно невысокой молекулярной массой порядка 24—29 тыс. Д и содержит в молекуле от 75 до 90 нуклеотидов. До 10 % всех нуклеотидов тРНК приходится на долю минорных оснований, что, по-видимому, защищает ее от действия гидролитических ферментов.
Роль тРНК заключается в том, что они переносят аминокислоты к рибосомам и участвуют в процессе синтеза белка. Каждая аминокислота присоединяется к определенной тРНК. Ряд аминокислот обладает более одной тРНК. К настоящему времени обнаружено более 60 тРНК, которые отличаются между собой первичной структурой (последовательностью оснований). Вторичная структура у всех тРНК представлена в виде клеверного листа с двухцепочным стеблем и тремя одноцепочными петлями (рис. 20). На конце одной из цепей находится акцепторный участок — триплет ЦЦА, к аденину которого присоединяется специфическая аминокислота. Аминокислота присоединяется к тРНК под действием фермента аминоацил-тРНК-синтетазы, который «узнает» одновременно и аминокислоту, и тРНК. В головке средней петли тРНК находится антикодон — триплет, состоящий из трех нуклеотидов. Антикодон комплементарен определенному кодону мРНК. При помощи антикодона тРНК «узнает» соответствующий кодон в иРНК, т. е. определяет место, куда
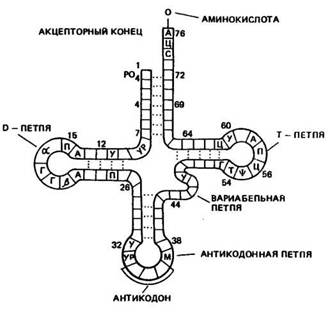
Рис, 20. Вторичная структура тРНК (по Ричу п Киму)
должна быть поставлена данная аминокислота в синтезируемой молекуле белка.
Предполагается, что петли тРНК, не вовлеченные в связывание и выполнение декодирующей функции аминокислоты, используются для связывания тРНК с рибосомой и со специфической аминоацил-тРНК-синтетазой.
Рибосомная РНК (рРНК). Размер рибосомных РНК эукариот составляет 5—28S (S — единица Сведберга, характеризующая скорость осаждения, седиментации частиц при ультрацентрифугировании), молекулярная масса 3,5-104— 1,510^ Д. Они содержат 120—3100 нуклеотидов. Рибосомная РНК накапливается в ядре, в ядрышках. В ядрышки из цитоплазмы транспортируются рибосомные белки, и там происходит спонтанное образование субчастиц рибосом путем объединения белков с соответствующими рРНК. Субчастицы рибосомы вместе или врозь транспортируются через поры ядерной мембраны в цитоплазму.
Рибосомы представляют собой органеллы величиной 20— 30 нм. Они построены из двух субчастиц разного размера и формы. На определенных стадиях белкового синтеза в клетке происходит разделение рибосом на субчастицы. Рибосомная РНК служит как бы каркасом рибосом и способствует первоначальному связыванию иРНК с рибосомой в процессе биосинтеза белка. Субчастицы обозначают у эукариот как 60 и 40S. Целые рибосомы
осаждаются при 80S. 40£-субчастица содержит 18S РНК и примерно 30 белков; бОБ-субчастица содержит 28S РНК, 5S РНК и 5,8 S РНК. В состав этой частицы входит примерно 50 различных белков. У прокариот функциональная рибосома имеет константу седиментации 70S. 708-рибосомы состоят из малой (30S) и большой (50S) субчастиц. 808-рибосомы содержат примерно равное количество рРНК и белка, у 70S-pn6ocoM соотношение РНК и белка составляет 2:1. Число рибосом в клетке прокариот равно примерно 104, у эукариот — около 105. В период синтеза белка рибосомы могут объединяться в полисомы, образуя более высокоорганизованные комплексы.

