 2014-02-09
2014-02-09 4002
4002Локализация нуклеиновых кислот в клетке
Четвертичная структура нуклеиновых кислот
Третичная структура нуклеиновых кислот
Молекулы ДНК в клетках живых организмов могут достигать нескольких сантиметров (при средней толщине до 10 нм). Они образуют плотную третичную структуру, толщина и длина которой не превышает нескольких нанометров. Это достигается путем суперспирализации, при которой двухцепочечные ДНК закручены в правую или левую суперспираль.
Третичная структура тРНК представляет собой глобулу, вытянутую в форме буквы «Г».
Четвертичная структура известная лишь для РНК. Она представляет собой блок из двух одинаковых субъединиц, связь между которыми поддерживается комплементарными взаимодействиями их азотистых соединений.
Локализация ДНК и РНК в клетке различна. ДНК локализована в ядре клетки, РНК – в ядре и цитоплазме. Только тРНК находится преимущественно к цитоплазме в растворенном виде. Как правило, нуклеиновые кислоты (ДНК и РНК) в живой клетке находятся в форме нуклеопротеинов. Основные структуры, содержащие нуклеопротеины, - это хроматин, где локализована ДНК, и рибосомы, где локализована РНК.
В клетке встречаются также полирибосомы – образования, состоящие из вытянутой молекулы мРНК, окруженной глобулярными белками, на которую нанизаны рибосомы.
К нуклеотидам, не входящим в состав ДНК и РНК, относятся нуклеотиды, встречающиеся в клетке или в свободном виде в качестве продуктов гидролиза или синтеза нуклеиновых кислот, или в виде компонентов многих важных соединений коферментной природы (моно- и динуклеотиды, содержащие один, два или три остатка фосфорной кислоты).
Одним из важнейших соединений нуклеотидной природы является аденозинтрифосфат (АТР) (рис. 4).
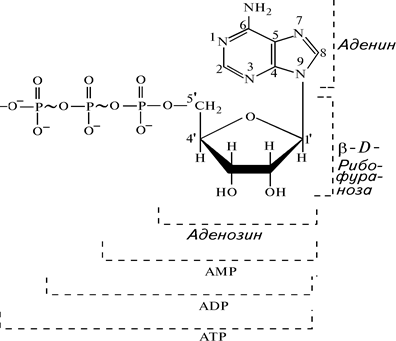
Рис. 4. Строение молекул АТР (аденозин-5'-трифосфата), АDР (аденозин-5'-дифосфата), АМР (аденозин-5'-монофосфата) и их компонентов
 АТР состоит из азотистого пуринового основания – аденина, D -рибозы и трех остатков фосфорной кислоты. Вторая и третья молекулы фосфорной кислоты присоединяются друг к другу макроэргической связью, которая при гидролизе освобождает большое количество химической энергии (от 40 до 60 кДж/моль) в зависимости от локализации реакции в клетке. Таким образом, в АТР имеются две макроэргические связи. АТР выступает в роли переносчика химической энергии, связывающей клеточные процессы, сопровождающиеся выделением энергии, с теми реакциями, которые протекают с потреблением энергии. Обратный процесс – синтез АТР из АDР и неорганического фосфора – сопровождается потреблением энергии:
АТР состоит из азотистого пуринового основания – аденина, D -рибозы и трех остатков фосфорной кислоты. Вторая и третья молекулы фосфорной кислоты присоединяются друг к другу макроэргической связью, которая при гидролизе освобождает большое количество химической энергии (от 40 до 60 кДж/моль) в зависимости от локализации реакции в клетке. Таким образом, в АТР имеются две макроэргические связи. АТР выступает в роли переносчика химической энергии, связывающей клеточные процессы, сопровождающиеся выделением энергии, с теми реакциями, которые протекают с потреблением энергии. Обратный процесс – синтез АТР из АDР и неорганического фосфора – сопровождается потреблением энергии:
 АDР + Р АТР
АDР + Р АТР
Подобная реакция называется реакцией фосфорилирования. Различают:
- Фотосинтетическое фосфорилирование (в этом процессе АТР синтезируется за счет солнечной энергии, которая используется при фотосинтезе).
- Гликолитическое фосфорилирование, когда синтез АТР происходит вследствие выделения энергии при окислении органических веществ без доступа кислорода воздуха (анаэробная фаза дыхания, или процесс гликолиза).
- Окислительное фосфорилирование, происходящее при окислении органических веществ при потреблении кислорода воздуха (аэробная фаза дыхания, или цикл трикарбоновых кислот, цикл лимонной кислоты, или цикл Кребса).
Другим соединением, у которого макроэргическая связь образуется между кислородом и фосфором, является UDP – уридиндифосфат. В состав UDP входит азотистое основание – урацил, D -рибоза и два остатка фосфорной кислоты, соединенные друг с другом макроэргической связью (рис. 5).
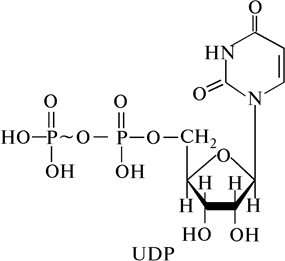
Рис. 5. Строение молекулы UDР (уридин-5¢-трифосфата)
UDP является коферментом гликозилтрансфераз, способных переносить глюкозу на какой-либо акцептор. Перенос глюкозы посредством UDP называется трансгликозилированием. UDP играет большую роль в превращении углеводов, в частности в синтезе сахарозы и крахмала.
Аналогичную роль переносчика молекул выполняет СТР – цитиндинтрифосфат (цитидинтрифосфорная кислота), участвующий в синтезе фосфолипидов – переносе остатков холина, этаноламина, серина и др. на фосфатидную кислоту.
Сложный нуклеозиддифосфат, называемый коферментом А (СоА), является переносчиком ацилов жирных кислот, которые в процессе синтеза связываются с концевой –SН-группой СоА макроэргической связью:
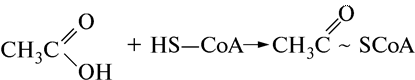
В обменных реакциях активироваться коферментом может не только уксусная кислота, но и другие органические кислоты. С образованием активированных остатков органических кислот протекают процессы аэробной фазы дыхания и распада жирных кислот и т.д.

