 2014-02-10
2014-02-10 5929
5929В зависимости от вида растений и условий их произрастания листья отличаются большим разнообразием. Однако можно выделить общие анатомические особенности, обеспечивающие возможность эффективного фотосинтеза.
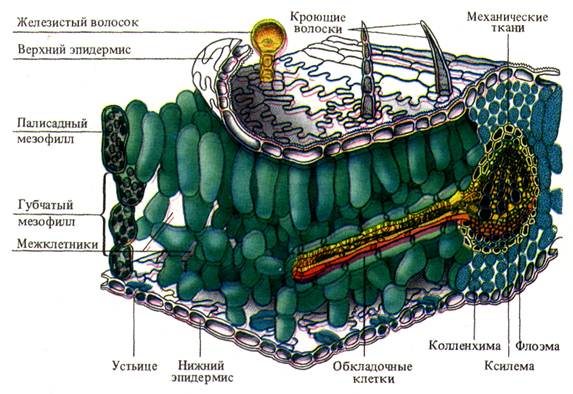
1. Наличие покровной ткани - эпидермиса, защищающего лист от излишней потери воды. Клетки нижнего и верхнего эпидермиса лишены хлоропластов, имеют крупные вакуоли, которые, подобно линзам, фокусируют свет на расположенную глубже хлорофиллоносную ткань. Деятельность клеток мезофилла основана не только на их хорошем освещении, она зависит также от поступления СО2. Нижний эпидермис, реже и верхний, имеют большое количество устьиц. Щели открытых устьиц занимают примерно 1 % площади листовой пластинки, диффузия СО2 внутрь листа идет через них сравнительно быстро. Отдельное устьице позволяет за 1 с поступить в лист 2500 млрд. молекул СО2. Поверхность листа поглощает СО2 только в 1,5-2 раза меньше, чем открытая поверхность щелей той же площади, хотя открытые устьица составляют лишь сотую часть поверхности. Такая высокая скорость связана с особенностями диффузии газов через мелкие отверстия, находящиеся на значительном расстоянии друг от друга, за счет краевого эффекта.
2. Наличие специализированной фотосинтетической ткани - хлоренхимы. Основная хлорофиллоносная ткань - палисадная паренхима - расположена обычно на освещаемой части листа. Вытянутость клеток и перпендикулярное расположение их к эпидермису обеспечивают увеличение поверхности, вдоль которой могут располагаться хлоропласты, не затеняя друг друга, а также облегчают отток ассимилятов. В каждой клетке полисадной паренхимы находится 30-40 хлоропластов. Губчатая ткань характеризуется меньшим содержанием хлоропластов (примерно 20 на клетку) и сильно развитой системой межклетников. Объем межклетников составляет 15-20 % общего объема листа и образует внутреннюю газовую среду, которая при помощи устьичных щелей сообщается с атмосферой. За счет межклетников значительно возрастает внутренняя рабочая поверхность, через которую каждой клеткой паренхимы поглощается СО2. Она в 8-12 раз больше, чем наружная поверхность листа. Хлоропласты как основные светоулавливающие органеллы еще в большей степени увеличивают светопоглощающую поверхность листа. На 1 см2 листа приходится примерно 200 см2 поверхности хлоропластов. Это является выражением общебиологической особенности организации - создание больших внутренних рабочих поверхностей при сравнительно малых наружных испаряющих площадях за счет, как было отмечено, затрат небольших количеств материала.
3. Наличие сильно развитой густой системы жилок - проводящих путей, что обеспечивает быстрый отток ассимилятов и снабжение фотосинтезирующих клеток водой и необходимыми минеральными веществами.
В зависимости от внешних условий, при которых происходят формирование и функционирование листьев, анатомическое строение их может существенно различаться. Листья, формирующиеся в условиях недостаточной влагообеспеченности, имеют ксероморфную структуру. В зависимости от освещения меняется соотношение между полисадной и губчатой паренхимой в мезофилле. Имеются и другие приспособления для функционирования листа в определенных условиях. Еще более существенные отклонения от типичного строения листа связаны с физиолого-биохимическими особенностями фиксации СО2 у С4-растений, к которым относятся кукуруза, сахарный тростник, ряд злостных сорняков наших полей.
Организация фотосинтетического аппарата на уровне листа может быть также охарактеризована на основе анализа его мезоструктуры. Понятие «мезоструктура» предложено А.Т. Мокроносовым в 1975 г. Оно охватывает целый ряд морфофизиологиических характеристик листа, позволяющих оценить ассимиляционную способность листа в целом. Основными показателями мезоструктуры листа являются: площадь листа, число клеток хлоренхимы на единицу площади листа, число хлоропластов в клетке и их объем, площадь поверхности хлоропластов, а также содержание хлорофилла в расчете на единицу площади листа, содержание ферментов углеродного цикла фотосинтеза в листе и их активность, общая интенсивность фотосинтеза. Показатели мезоструктуры листа могут значительно варьировать в зависимости от внешних факторов среды, а также от физиологического состояния растений.
Лист представляет собой уникальную оптическую систему - ловушку для света. Как видно из рисунка, лист поглощает значительную часть излучения в видимой области спектра (400-700 нм). Большую часть синего и красного света поглощают пигменты хлоропластов первых слоев клеток хлоренхимы. Свет, не поглощенный в верхних слоях листа, обогащен лучами зеленой области спектра. Его многократное отражение от стенок мезофильных клеток в толще листа значительно увеличивает оптический путь и повышает вероятность поглощения лучей этой области спектра, несмотря на то что коэффициент поглощения хлорофилла в зеленой области значительно меньше, чем в синей и красной.
Структура листа обеспечивает поступление углекислого газа из воздуха для реакций фотосинтетической ассимиляции углерода. Газообмен мезофилла листа с окружающим воздухом осуществляется через устьица. Количество устьиц, их свойства (величина устьичной щели, устьичное сопротивление и др.) являются важными факторами регуляции фотосинтеза. Значительный объем подустьичного пространства, а также достаточно рыхлое расположение мезофильных клеток в листе и существование больших межклетников в ткани хлоренхимы позволяют создать большие воздушные запасы в толще листа для непрерывного снабжения углекислым газом углеродных циклов фотосинтеза.
Лист - донор ассимилятов в растении. Отток органических веществ, образовавшихся в процессе фотосинтеза, происходит по клеткам сосудистых пучков флоэмы. Транспорт ассимилятов из автотрофных клеток предполагает вынос сахарозы (основного транспортного соединения фотосинтеза) из авто-трофной клетки в апопласт и последующую загрузку флоэмы листа.

