 2014-02-12
2014-02-12 1004
1004Т-лимфоциты, растворимые цитокины и макрофаги – от внутриклеточных.
Антитела, комплемент и полиморфоядерные лейкоциты обеспечивают защиту от большинства внеклкточных чужеродных агентов, а
Природа узнавания антигена антителом определяется аминокислотной последовательностью гипервариабельных участков молекул антител.
В различных антителах даже те области молекулы, аминокислотная последовательность которых не варьирует (константные) не все одинаковы.
У млекопитающих константные области легких цепей бывают двух типов æ и l (95:5 у мыши и 70:30 у человека). Константные области тяжёлых цепей, существуют пяти классов: m,d,g,e,a. Антитела, в которых вариабельной домен одинаковый, но тяжелые цепи разных классов, узнают один и тот же антиген, но играют разную роль в иммунном ответе. Например, связанные с мембраной антитела, служащие рецепторами В-клеток, принадлежат по типу тяжелых цепей к классу m или d, а большая часть секретируемых антител – к классу g или a.
Тимус представляет собой орган, принимающий Т-клетки и осуществляющий созревание или отбор антигенспецифических Т-клеток и осуществляющий избирательное освобождение этих клеток на периферию.
Процесс созревания сопровождается «включением» специфических генов, причем некоторые из них экспессируются только во время той фазы дифференцировки, которая происходит в тимусе, тогда как другие остаются в активном состоянии на протяжении всей жизни Т-клетки.
На определенных стадиях созревания тимоцитов происходят следующие события:
экспрессия антиген-специфических рецепторов;
появление или отбор клеток, распознающих продукты специфических для тимуса МНС-генов, необходимых для взаимодействия клеток иммунной системы;
появление на клетках достаточного количества рецепторов, отвечающих за доставку лимфоцитов из кровяного русла в организованные лимфоидные ткани;
приобретение Т-клетками способности осуществлять помощь, «убийство» или супрессию.
Механизм созревания и последовательность стадий созревания пока не ясны.
Зрелый тимус представляет собой эпителиально-лимфоидный орган, состоящий из трех отдельных слоев (наружного и внутреннего коркового и мозгового), каждый из которых содержит лимфиидные клетки одного определенного класса и нелимфоидные клетки, обеспечивающие необходимое микроокружение для созревания лимфоцитов.
В наружном субкапсулярном корковом слое обнаруживаются примитивные делящиеся клетки (лимфобласты). Некоторые из них, очевидно, взаимодействуют со специализированными эпителиальными клетками-кормилицами, ускоряющими процессы самообновления (пролиферации) лимфобластов и превращения их во все другие тимусные клетки и Т-клетки различных подклассов.
На долю крупных самообновляющихся тимусных лимфобластов приходится 5-15% общего количества тимусных лимфоцитов. Они дают начало трем популяциям клеток:
малым тимоцитам внутреннего коркового слоя,
некоторым тимоцитам среднего размера, расположенным в мозговом слое,
тимусным клеткам эмигрантам.
Сами лимфобласты образуются, по-видимому, из заселяющих тимус предшественников после длительного лаг-периода, в течении которого эти предшественники дифференцируются и мигрируют в наружный корковый слой тимуса.
Регуляция процессов созревания тимусных лимфоцитов и их размещения в тимусе частично связана с избирательной экспрессией специфических гликозилтрансфераз или гликозидаз.
Во внутреннем корковом слое находятся главным образом потомки лимфобластов – в большинстве своем неделящиеся малые тимусные лимфоциты, располагающиесявблизи дендритных корковых эпителиальных клеток.
Внутренний корковый слой тимуса содержит основную часть тимусных лимфоцитов, большинство из которых погибает, и особый класс дендритных эпителиальных клеток.
Малые тимоциты составляющие примерно 85% общего числа всех тимусных лимфоцитов, обладают in vivo, подобно другим корковым лимфоцитам, ярко выраженной чувствительностью к кортизону.
Интересно, что в популяции малых тимоцитов внутреннего коркового слоя синтезируется большое количество внутриклеточной терминальной дезоксинуклеотидилтрансферазы и в клетках данной популяции постоянно поддерживается высокая концентрация фермента.
Возможно, гибель значительной части этих клеток в тимусе обусловлена в той или иной мере либо продолжительным действием терминальной дезоксинуклеотидилтрансферазы, либо гибель этих клеток может быть обусловлена их высокой чувствительностью к гликокортикоидам эндогенного происхождения.
Не исключено, что немногие зрелые или подвергшиеся соответственному отбору клетки преобретают способность покидать тимус только после завершения гликозилирования их поверхностных гликопротеинов и гликолипидов.
Сформировавшиеся тимусные лимфоциты среднего размера движутся по направлению к мозговому слою – на их пути расположены корковые макрофаги, выполняющие функцию разрушителей («могильщиков»). Они участвуют в разрушении и фагоцитозе уже погибших или обреченных на гибель тимусных лимфоцитов.
Некоторое количество пре-Т-клеток содержится в костном мозге, они представляют собой отдельную популяцию, хотя, по-видимому, имеющую общего предшественника с пре-В-клетками.
Процессы созревания В-лимфоцитов в костном мозге и Т-лимфоцитов в тимусе имеют важную общую особенность: в первичных лимфатических органах производится гораздо больше клеток, чем их обнаруживается в общем пуле периферических иммунокомпетентных лимфоцитов.
Итак, из стволовой клетки костного мозга или из пре-Тклетки в тимусе созревают и дифференцируются три популяции Т-клеток:
ТE – Т-эффекторы (киллеры);
Тh - Т-хелперы (помощники);
TS - Т-супрессоры.
Кроме Т- и В- клеток в периферической крови примерно 10-20% лимфоцитов не несут отличительных маркеров Т- или В-лимфоцитов – нулевые клетки.
Тимоциты периферической крови различаются тропностью: одна субпопуляция (Т1) оседает в селезенке, другая (Т2) – в лимфатических узлах.
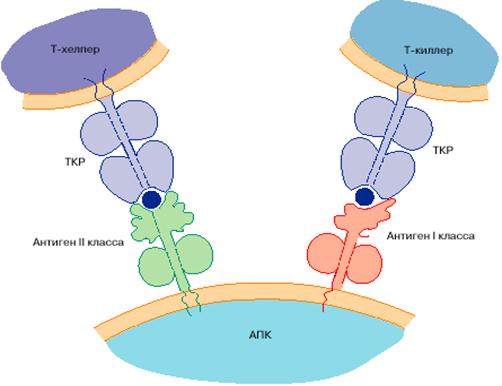
Т-эффекторные клетки или киллеры (цитотоксические Т-клетки) – они лизируют клетки, несущий на своей поверхности те антигены, к которым специфичны данные лимфоциты. Лизис клетки-мишени ТЕ –лимфоцитами – процесс сложный, проходит в несколько стадий. Первая стадия – распознавание соответствующего антигена и молекул класса I, имеющихся на поверхности клетки-мишени. Вторая стадия – программирование для лизиса, ТЕ-клетка вызывает повреждение клетки-мишени, а затем следует ее лизис. Отдельная ТЕ-клетка может участвовать в лизисе нескольких клеток по очереди, сама при этом не разрушаясь.
Т-хелперы (to help – помогать) и Т-супрессоры (to supress – подавлять) выполняют регуляторные функции. Т-хелперы, узнав антиген, стимулируют остальные компоненты иммунной системы, то есть В-клетки и другие Т-клетки, специфичные к данному антигену.
Т-хелперы способны стимулировать В-клетки к пролиферации и дифференцировке в антителообразующие клетки, причем, обнаружено несколько механизмов такого стимулирования:
-прямое взаимодействие ТН-клетки и реагирующей В-клетки (когнатная помощь). Природа медиатора, передаваемого от Т-клетки к В-клетке при когнатной помощи пока не установлена;
-путем образования растворимых неспецифических хелперных факторов, часто называемых лимфокинами;
- когнатное взаимодействие плюс медиатор.
|
Название «Т-хелперы» создает впечатление, что они играют подчиненную, или второстепенную, роль, как-будто эти клетки только поощряют процессы, которые проходили бы и без них. На самом деле они, по-видимому, являются «главным выключателем» иммунной системы. В-клетки узнают антиген независимо от Т-клеточной стимуляции, но их пролиферация и окончательная дифференцировка требуют активации Т-хелперами.








