 2014-02-12
2014-02-12 5534
5534Нуклеиновые кислоты играют очень важную роль в жизнедеятельности организмов и наряду с белками определяют главнейшие звенья обмена веществ, явления роста и размножения организмов, а также передачу наследственной информации. Содержание ДНК на одну клетку довольно постоянно. В органах и тканях, характеризующихся быстрым ростом, интенсивно идет клеточное деление, значит, с такой же интенсивностью идет новообразование, синтез новых молекул ДНК. С другой стороны, в старых органах и тканях, характеризующихся интенсивным отмиранием клеток, преобладает распад молекул дезоксирибонуклеиновой кислоты. Содержание РНК в среднем на 1 клетку не остается постоянным в течение всего жизненного цикла клетки. Сразу после клеточного деления оно невысоко, но быстро увеличивается и достигает максимума в период наиболее интенсивного роста клетки, когда напряженность обмена веществ, и прежде всего интенсивность синтеза белков, в клетке бывает самой высокой. В дальнейшем содержание РНК в клетке снижается, происходит ее распад. Таким образом, простые определения содержания ДНК и РНК в клетках и тканях показывают, что в организмах постоянно идут процессы синтеза и распада нуклеиновых кислот.
Биосинтез нуклеиновых кислот. Основными структурными элементами, из которых построены молекулы ДНК и РНК, являются нуклеотиды – соединения, состоящие из рибозы и дезоксирибозы, азотистого основания и фосфорной кислоты. Поэтому образованию нуклеиновых кислот в клетках должен, очевидно, предшествовать синтез нуклеотидов. На второй стадии биосинтеза происходит фосфорилирование нуклеотидов с образованием соответствующих ди- и трифосфатов, имеющих макроэргические фосфатные связи. На третьей, последней стадии синтеза нуклеиновых кислот соответствующие фосфорилированные нуклеотиды полимеризуются, и возникают молекулы ДНК и РНК. Все животные организмы способны синтезировать нуклеиновые кислоты из простых соединений и не нуждаются в доставке их с пищей. Хорошим примером этому служит рост молодых животных, которые питаются только молоком.
Биосинтез пуриновых нуклеотидов. В отличие от многих других синтетических процессов, биосинтез пуриновых нуклеотидов происходит чаще всего не при взаимодействии соответствующих веществ, входящих в состав этих нуклеотидов (пуриновых оснований, рибозы, дезоксирибозы и фосфорной кислоты), а в результате более сложных реакций. Оказалось, что исходным соединением при биосинтезе нуклеотидов является рибоза, или рибозо-5-фосфат. К рибозо-5-фосфату затем последовательно присоединяются отдельные атомы или группы атомов, из которых постепенно строится гетероциклический скелет пуринового основания. В образовании этого скелета принимают участие атомы азота и углерода ряда аминокислот, муравьиной кислоты и угольной кислоты. Процесс требует значительной затраты энергии. Донором формильных остатков (остатков муравьиного альдегида) является производное фолиевой кислоты – ангидроформилтетрагидрофолиевая кислота (АФТГФК). Синтез АФТГФК происходит из тетрагидрофолиевой кислоты (ТГФК) с участием АТФ по схеме:
 НСООН+ТГФК+АТФ АФТГФК+АДФ+Н3РО4.
НСООН+ТГФК+АТФ АФТГФК+АДФ+Н3РО4.
Промежуточным продуктом в синтезе пуриновых нуклеотидов является инозиновая кислота. Она состоит из пуринового основания – гипоксантина, рибозы и фосфорной кислоты. Пуриновые нуклеотиды – адениловая, дезоксиадениловая, гуаниловая и дезоксигуаниловая кислоты – образуются из инозиновой кислоты в результате довольно простых превращений. Рибозо-5-фосфат, необходимый для синтеза нуклеотидов, образуется в растениях в процессе фотосинтеза или при окислении углеводов через пентозофосфатный цикл. Процесс биосинтеза инозиновой кислоты довольно сложен и идет через несколько стадий.
Суммарное уравнение биосинтеза инозиновой кислоты из простейших предшественников можно схематически представить следующим образом:
 3 NH4+ + 2 HCOO– + HCO3– + глицин + рибозо-5-фосфат инозиновая кислота + 9 Н2О.
3 NH4+ + 2 HCOO– + HCO3– + глицин + рибозо-5-фосфат инозиновая кислота + 9 Н2О.
При этом две молекулы аммиака отщепляются от глютамина и одна – от аспарагиновой кислоты, а остатки муравьиной кислоты переносятся от 2 молекул ангидроформилтетрагидрофолиевой кислоты. Всего для синтеза 1 молекулы инозиновой кислоты из рибозо-5-фосфата необходимо 5 молекул АТФ. Но при расчете общего количества энергии следует учитывать, что при образовании каждой молекулы инозиновой кислоты необходимо девять молекул АТФ. Синтез адениловой кислоты из инозиновой кислоты идет в две стадии через аденил-янтарную кислоту. Донором атома азота является аспарагиновая кислота. Эта реакция во многом напоминает реакцию синтеза инозиновой кислоты. Основное различие состоит в используемых макроэргических соединениях, которые дают энергию для синтеза С-N-связи. В реакции участвует АТФ, а в реакции синтеза адениловой кислоты – гуанозинтрифосфат (ГТФ). На 2-й стадии от аденилянтарной кислоты отщепляется фумаровая кислота, и образуется адениловая кислота. Обе реакции синтеза адениловой кислоты катализируются ферментом аденилосукцинатлиазой. Биосинтез гуаниловой кислоты из инозиновой кислоты также идет в две стадии и требует затрат энергии. На 1-й стадии инозиновая кислота окисляется до ксантиловой кислоты при помощи восстановленного никотинамидадениндинуклеотида. Эта реакция катализируется ферментом инозин-5-фосфатдегидрогеназой. На 2-й стадии происходит аминирование ксантиловой кислоты, которое требует затрат энергии. Донором атома азота здесь является глутамин. В результате реакции образуется гуаниловая кислота, аденозинмонофосфат и пирофосфорная кислота. Пуриновые дезоксирибонуклеотиды – дезоксиадениловая и дезоксигуаниловая кислоты, которые необходимы для биосинтеза ДНК, – образуются аналогично. Исходным продуктом для их синтеза также является рибоза, а превращение производных рибозы в производные дезоксирибозы осуществляется в результате их восстановления на уровне нуклеотидов при сохранении гликозидной связи между пентозой и основанием. Для восстановления необходим НАДФ*Н2.
Биосинтез пиримидиновых нуклеотидов. Механизм биосинтеза пиримидиновых нуклеотидов, так же как и пуриновых нуклеотидов, был выяснен в основном лишь в последние годы. Хотя по своей структуре пуриновые и пиримидиновые нуклеотиды довольно близки между собой, тем не менее пути их биосинтеза резко различны. Если при биосинтезе пуриновых нуклеотидов исходным соединением является рибозо-5-фосфат, к которому постепенно присоединяются отдельные атомные группировки, то биосинтез пиримидиновых нуклеотидов начинается с самых простых соединений (аммиака и углекислоты), а рибофосфат присоединяется лишь на заключительных стадиях синтеза. Промежуточным продуктом в синтезе производных пиримидина является оротовая кислота.
Но в результате этих реакций синтез нуклеотидов происходит не всегда. В тех случаях, когда в тканях и клетках растений имеются свободные пуриновые и пиримидиновые основания (которые возникают в результате распада нуклеиновых кислот), они могут непосредственно использоваться для синтеза нуклеотидов. При синтезе из свободных оснований вначале образуются нуклеозиды, которые затем фосфорилируются и превращаются в нуклеотиды.
На первом этапе под действием фосфотрансфераз фосфатный остаток переносится от АТФ на рибозу или дезоксирибозу с образованием пентозофосфатов. Затем пентозофосфат, соединяясь с основанием, превращается в нуклеотид и свободную фосфорную кислоту. На последнем этапе нуклеотид фосфорилируется с участием АТФ и превращается в нуклеотид. Схематически процесс биосинтеза нуклеотида из свободного основания можно представить следующим образом (на примере синтеза адениловой кислоты):
 рибозо-1-фосфат + аденин аденозин + Н3РО4
рибозо-1-фосфат + аденин аденозин + Н3РО4
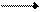 аденозин + АТФ адениловая кислота + АДФ
аденозин + АТФ адениловая кислота + АДФ
С интез ДНК. Химические анализы показывают, что количество ДНК в течение жизненного цикла клетки постоянно. Но в период клеточного деления количество ДНК резко возрастает, и ее концентрация увеличивается ровно в два раза. Таким образом, после деления клетки содержание ДНК в дочерних клетках остается таким же, каким оно было в материнской клетке.
Молекула ДНК представляет собой две комплементарные полинуклеотидные цепи, скрученные вокруг общей оси, причем в цепях аденин соответствует тимину, а гуанин – цитозину, соединенным водородными связями. Таким образом, каждая цепь ДНК служит специфической структурой, которая может соединяться только с комплементарной структурной цепью и точно определять структуру вновь создаваемой цепи. В процессе удвоения молекул ДНК в период клеточного деления прежде всего разрываются водородные связи между цепями, цепи раскручиваются и расходятся. После этого под действием соответствующих ферментов к каждой из одиночных цепей присоединяются новые нуклеотиды. Но так как сочетание должно быть строго определенным, то на каждой образовавшейся цепи строится вторая комплементарная цепь прежнего состава.
Этот процесс удвоения ДНК представлен на схеме:


 –А–Ц–Т–Т–Г–
–А–Ц–Т–Т–Г–



 –А–Ц–Т–Т–Г– –А–Ц–Т–Т–Г–
–А–Ц–Т–Т–Г– –А–Ц–Т–Т–Г–


 –Т–Г–А–А–Ц–
–Т–Г–А–А–Ц–

 –Т–Г–А–А–Ц–
–Т–Г–А–А–Ц–
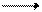 –Т–Г–А–А–Ц– –Т–Г–А–А–Ц–
–Т–Г–А–А–Ц– –Т–Г–А–А–Ц–
–А–Ц–Т–Т–Г–
Таким образом, благодаря принципу комплементарности в строении молекулы из одной молекулы ДНК образуются две совершенно одинаковые молекулы. Способность ДНК давать строго определенные, подобные себе новые молекулы играет определяющую роль в явлениях наследственности и в передаче генетической информации. ДНК определяет синтез специфических белков в клетке, и изменения в ее структуре будут вести к синтезу неспецифических для данной клетки и организма белков, что в конечном итоге вызывает изменения в обмене веществ и свойствах организма. Поэтому сама ДНК должна сохранять постоянство строения, не изменять его даже при делении клеток, что и достигается в результате специфического механизма самоудвоения ее молекулы. В периоды между делениями клеток молекула ДНК остается очень инертной, стабильной, что резко отличает ее от всех других веществ – белков, углеводов, липидов, которые подвергаются непрерывному обмену и обновлению. Одним из наиболее крупных достижений биохимии явилось выделение и очистка ферментов, катализирующих синтез нуклеиновых кислот. Эти работы позволили воспроизвести синтез ДНК и РНК вне живой клетки. В 1956 году А. Корнберг выделил и очистил из экстрактов кишечной палочки фермент, катализирующий синтез ДНК из ее простых предшественников. Фермент был назван ДНК-полимеразой, а впоследствии получил название – ДНК-нуклеотидилтрансфераза. Позднее этот фермент был выделен из других объектов. Он катализирует синтез ДНК только из соответствующих трифосфатов, при замене трифосфатов на дифосфаты или монофосфаты синтеза ДНК не было. Необходимым условием для выявления максимальной активности ДНК-нуклеотидилтрансферазы является одновременное присутствие в среде трифосфатов всех четырех дезоксирибонуклеозидов (дезокситимидина, дезоксицитидина, дезоксигуанозина, дезоксиаденозина), а также ионов магния и небольшого количества ДНК (так называемой затравки). Суммарную реакцию синтеза ДНК можно представить следующим образом:
ng АТФ



 + g АФ
+ g АФ
ng ЦТФ полимераза
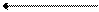
 + + ДНК ДНК – g ЦФ + 4 n ФФ,
+ + ДНК ДНК – g ЦФ + 4 n ФФ,
ng ГТФ g ГФ
+
 ng ТТФ g ТФ n
ng ТТФ g ТФ n
где g АТФ, g ЦТФ, g ГТФ, g ТТФ – соответственно дезоксиаденозинтрифосфат, дезоксицитидинтрифосфат, дезоксигуанозинтрифосфат и дезокситимидинтрифосфат, g АФ, g ЦФ, g ГФ, g ТФ – соответствующие нуклеотиды, соединенные в цепь, ФФ – неорганический пирофосфат. Эта реакция обратима, и процесс идет в сторону синтеза при условии выделения из среды неорганического пирофосфата. Состав ДНК в процессе ее биосинтеза обычно не меняется, однако в некоторых случаях изменения все же происходят. Например, в силу каких-то причин имеющиеся основания могут быть заменены их аналогами или вовсе выпасть из молекулы. Такие изменения состава ДНК носят название «случайных ошибок» биосинтеза ДНК. Совершенно очевидно, что если в результате «случайных ошибок» образуется ДНК не характерная для данного вида, то она после самоудвоения дает начало таким же не характерным молекулам, что приводит к спонтанным мутациям организма. Но такие «случайные ошибки» встречаются исключительно редко. Однако часто мутации можно увеличить искусственно, если подвергнуть организм действию ионизирующей радиации, ультрафиолетовых лучей или химических веществ под названием мутагены. К мутагенам относят некоторые аналоги пуриновых и пиримидиновых оснований, алкирующие агенты – иприт, некоторые антибиотики, алкалоиды, азотистую кислоту.
Синтез РНК. Содержание РНК во всех клетках не постоянно, а может резко изменяться в зависимости от вида, возраста и физиологического состояния организма или ткани. Обычно количество РНК больше в молодых клетках, через в старых. Кроме того, в клетках содержатся различные по свойствам и молекулярному весу виды РНК: рибосомная, транспортная и информационная. Все это создает значительные трудности при изучении проблемы биосинтеза рибонуклеиновых кислот. В настоящее время считают, что биосинтез рибонуклеиновых кислот может происходить в результате одного из следующих типов реакций:
· действия полинуклеотидфосфорилазы;
· наращивания имеющихся полинуклеотидных цепей;
· ДНК–зависимого синтеза РНК;
· РНК–зависимого синтеза РНК.
1.Синтез РНК из рибонуклеозиддифосфатов катализирует фермент полинуклеотидфосфорилаза. Для действия фермента необходимо наличие в реакционной среде ионов магния, при синтезе РНК освобождается неорганический фосфат. Реакция синтеза РНК идет по схеме:
+ n Н2О
 n [рибонуклеозиддифосфат] n [рибонуклеотид РНК] + n Н3РО4
n [рибонуклеозиддифосфат] n [рибонуклеотид РНК] + n Н3РО4
 – n Н2О
– n Н2О
В качестве источников дифосфатов могут служить АДФ, УДФ, ГДФ и ЦДФ. Для синтеза высокомолекулярных полимеров в реакционную смесь необходимо добавлять некоторое количество «затравки». Состав «затравки» не оказывает влияния на состав синтезируемого продукта. В качестве «затравки» можно взять любую РНК или любой полинуклеотид. Состав синтезирующейся РНК зависит только от количества и соотношения нуклеозиддифосфатов в реакционной среде. Если в среде будет преобладать какой-нибудь нуклеозиддифосфат, то он же преобладает в составе РНК.

